Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/
|
|
|
- Olga Sedláčková
- před 8 lety
- Počet zobrazení:
Transkript
1 Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/

2 Molekulární markery PCR, RAPD, RFLP, AFLP, mikrosatelity, sekvenace

3 Genetické markery Genetické markery jsou definovány jako morfologické, anatomické, fyziologické nebo biochemicko-genetické vlastnosti organismů těsně korelující s jinými, mnohdy složitějšími, vlastnostmi.

4 Rozdělení genetických markerů morfologicko-anatomické biochemicko-fyziologické markery zásobní a stavební proteiny biochemicko-genetické izoenzymy nukleové kyseliny

5 Základní rozdělení DNA markerů délkového polymorfismu fragmentů DNA vzniklých restrikčním štěpením (RFLP) detekce variability na základě amplifikace fragmentů DNA v in vitro podmínkách (PCR) různých typů propojení metod RFLP a PCR (AFLP) polymorfismu repetetivních sekvencí mini- a mikrosatelitů stanovení sekvencí neboli pořadí nukleotidových párů v molekule DNA

6 Oblasti využití genetických markerů humánní medicína veterinární medicína kriminalistika šlechtění a plemenářství evoluční biologie paleontologie detekce letálně působících genů detekce napadení člověka například viry určení paternity detekce napadení zvířat například viry detekce letálně působících genů jednoznačná identifikace osob selekce s využitím markerů identifikace odrůdové pravosti a čistoty určení paternity fytopatologie historie

7 Selekce s využitím markerů Selekce s využitím markerů neboli MAS (Marker Assisted Selection) je aktuální trend ve šlechtění rostlin i zvířat. MAS je založeno na tom, že šlechtitel selektuje potomstvo na základě genetické analýzy s využitím různých typů markerů. Tímto způsobem eliminuje chybu selekce u znaků, které jsou modifikovány vnějším prostředím. Použití metody MAS výrazně urychluje a zefektivňuje šlechtění. Řada genetických markerů vykazuje kodominanci a umožňují tak odlišit homozygoty od heterozygotů. U kvantitativních znaků jsou pomocí MAS identifikovány takzvané kandidátní geny, které vykazují majoritní efekt na úroveň kvantitativního znaku. DNA markers can be used to determine whether a seedling carries an allele of interest. Here, the marker for the red-flesh allele is detected as an additional DNA band on an agarose gel. Breeders use information like this to decide which seedlings to grow and which to discard.

8 Biochemicko-genetické markery - detekce variability organismů přímo na úrovni jejich genetické informace. - starší typy markerů (proteiny, izoenzymy) detekovaly variabilitu produktů transkripce a translace. - současnost: detekce variabilitu nositelů genetické informace DNA a RNA. - zpravidla je variabilita detekována a vyhodnocována na základě elektroforetické separace různě velkých molekul obvykle na gelovém nosiči.

9 Markery založené na polymorfismu DNA Základní vlastnosti DNA markerů: jsou aplikovatelné u všech organismů, kde je zvládnutá technika izolace DNA nejsou závislé na podmínkách prostředí jejich počet je téměř neomezený často lze DNA markery lze použít na minimálním množství DNA = nedestruktivní metody (části listů, semena) pomocí DNA markerů lze charakterizovat i velmi raná ontogenetická stádia organismů
 pomocí DNA markerů lze charakterizovat i velmi raná ontogenetická stádia")

 (potenciální zdroj genů tolerance pšenice vůči salinitě a suchu).")
10 Proteinové markery Jsou známy vztahy mezi přítomností určité bílkoviny, lokalizací jejího genu a dalšími vlastnostmi rostliny. Pomocí těchto markerů je možné predikovat například kvalitu složení obilky, odolnosti k některým chorobám nebo stanovit odrůdovou pravost vypracovány certifikované metodiky U skotu je obdobným způsobem provedeno hodnocení proteinů mléka. Příklad využití proteinových markerů při studiu linií pýru (Agropyron elongatum) (potenciální zdroj genů tolerance pšenice vůči salinitě a suchu). Přítomnost specifických bandů u linií s vlastnostmi, které by mohly ovlivňovat vlastnosti těsta.
 (potenciální zdroj genů tolerance pšenice vůči salinitě a suchu).")
11 Isozymy = allozymy U většiny enzymů existují takzvané izoformy neboli izoenzymy. Izoenzymy katalyzují stejnou biochemickou reakci, t.j. reagují s jedním substrátem a vytváří shodný produkt. Izoformy jednoho enzymu se vzájemně odlišují například velikostí molekuly. Levné, rychlé, dostupné? Omezená hodnota informací Exprese závislá na prostředí Jen čerstvý materiál

12 Isozymy = allozymy


13 Isozymy = allozymy

14 RFLP RFLP - Restriction Fragment Lenght Polymorphism Polymorfismus v délce restrikčních fragmentů gdna naštěpíme restriktázou fragmenty rozdělíme elektroforeticky z gelu je přesajeme na membránu membránu hybridizujeme se sondou navázanou sondu detekujeme U malých genomů nebo malých fragmentů DNA s malým počtem vznikajících fragmentů metoda využívá pouhé elektroforetické separace různě dlouhých fragmentů U větších genomů technika využívá DNA-DNA hybridizace na principu Southern blottingu.


15 Co jsou to restrikční fragmenty? Restrikční fragmenty jsou úseky molekul DNA, které vznikají enzymatickou aktivitou restrikčních endonukleáz, které rozpoznávají specifické sekvence, kde přestřihnou molekulu DNA.

16 Postup RFLP Rozpoznávací místo restriktázy EcoRI - 6bp dlouhou sekvence (GAATTC) ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC Sonda -Ze všech naštípaných fragmentů vybírám pouze určitou část DNA, kterou studujeme (gen)

17 Postup RFLP Rozpoznávací místo restriktázy EcoRI - 6bp dlouhou sekvence (GAATTC) ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC
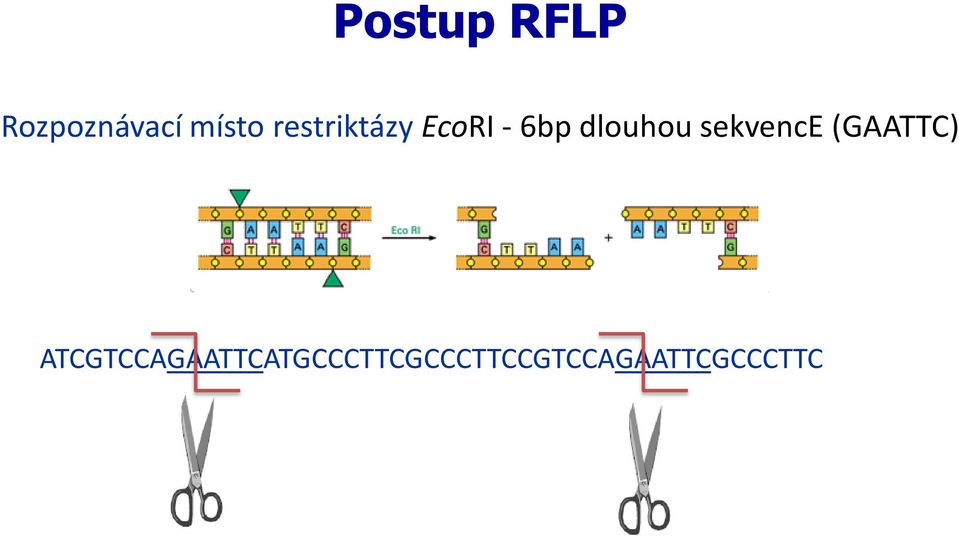
18 Postup RFLP Standartní (popř. dominantní) alela po naštěpení ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC ATCGTCCAG AATTCATGCCCTTCGCCCTTCCGTCCAG AATTCGCCCTTC
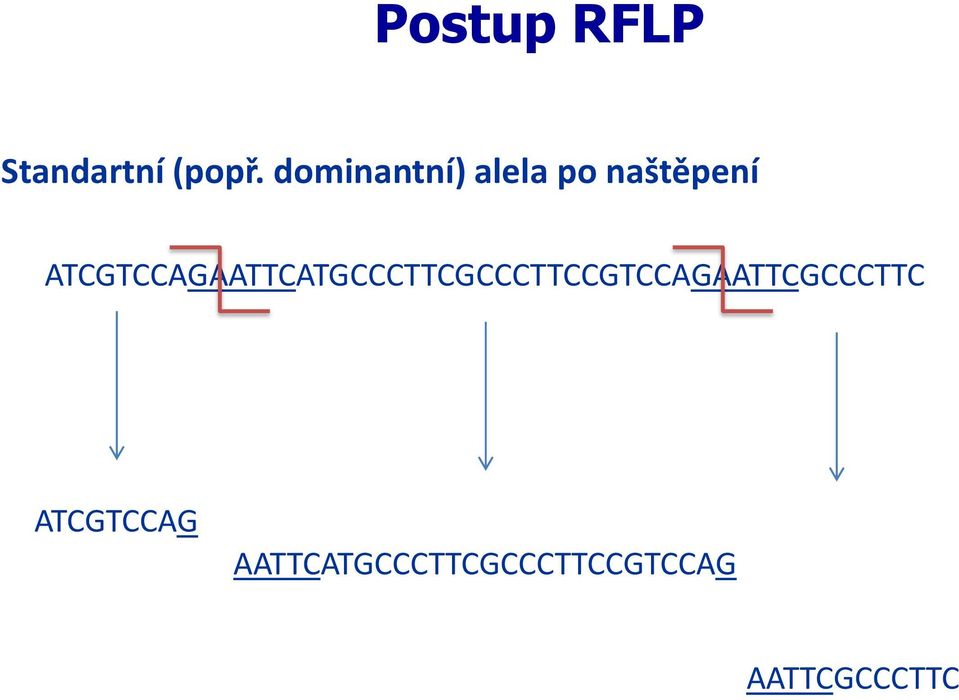
 alela po naštěpení ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC")
19 Postup RFLP Standartní (popř. dominantní) alela po naštěpení ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC ATCGTCCAG AATTCATGCCCTTCGCCCTTCCGTCCAG AATTCGCCCTTC Sonda
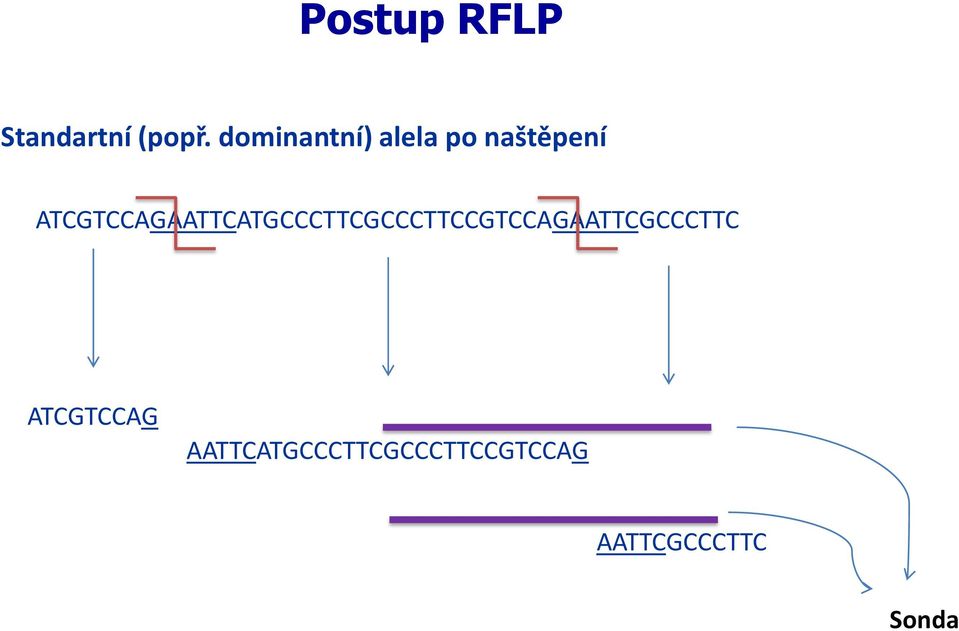
20 Postup RFLP Mutovaná (popř. recesivní) alela ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGACTTCGCCCTTC ztráta restrikčního místa (nezabrání vazbě sondy)
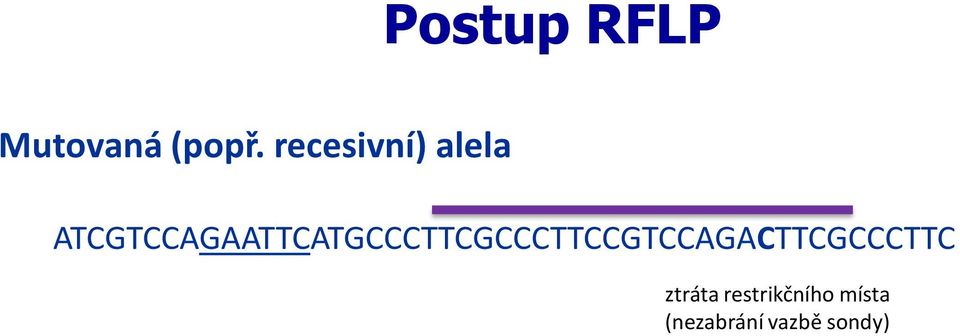
21 Postup RFLP Mutovaná (popř. recesivní) alela po naštěpení ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGACTTCGCCCTTC ATCGTCCAG AATTCATGCCCTTCGCCCTTCCGTCCAGACTTCGCCCTTC ztráta restrikčního místa (nezabrání vazbě sondy)
22 Postup RFLP Standartní alela Mutovaná alela ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC ATCGTCCAGAATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC Štěpení a elfo AATTCATGCCCTTCGCCCTTCCGTCCAG AATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC AATTCGCCCTTC ATCGTCCAG ATCGTCCAG Po provedení elektroforézy, přesátí a hybridizace (Southern blotting) AATTCATGCCCTTCGCCCTTCCGTCCAG AATTCATGCCCTTCGCCCTTCCGTCCAGAATTCGCCCTTC AATTCGCCCTTC ATCGTCCAG ATCGTCCAG
23 RFLP Sondou se rozumí úsek jednořetězcové DNA, který má sekvenci jako detekovaný gen. Sonda je obvykle označena radioizotopem fosforu, který ji umožňuje detekovat. classes.midlandstech.edu
24 Genetická interpretace RFLP markerů RFLP markery představují kodominantní typ markerů. Umožňují vzájemně odlišit dominantní homozygoty, heterozygoty a recesivní homozygoty. Uplatňují se například: 1. Pro vyhledávání v genomových knihovnách 2. Pro detekce konkrétních genů 3. Pro detekce bodových mutací 4. Pro tvorbu RFLP map,.

25 Segregace RFLP markeru Letální recesivní alela vznikla mutací a liší se od dominantní alely ztrátou restrikčního místa.
26 Případ Colin Pitchfork (1983 a 1986) Dvě slečny znásilněny a zavražděny v hrabství Leicestershire v Anglii Mentálně zaostalý mladík Richard Buckland, který nahlásil nález druhého těla, se přiznal k zavraždění této dívky Na základě RFLP vyloučen místních mužů bylo požádáno o vzorek krve nebo bukální stěr Pekař exhibicionista Colin Pitchfork za sebe k odběru poslal (za 200 liber) kamaráda Iana Kellyho, který se tím pochlubil v hospodě První omilostnění a první obvinění pomocí DNA důkazů.
27 Nevýhody RFLP Požadavek velkého množství intaktní DNA (50 ng odpovídá ~ buněk) Metoda málo robustní (interpretační potíže)
28 Replikace in vivo
29 Amplifikace fragmentů DNA v in vitro podmínkách Tato metoda detekce genů je označována jako polymerázová řetězová reakce PCR (Polymerase Chain Reaction). Je založena na zmnožení hledaného úseku DNA. Který je ohraničen známými sekvencemi takzvanými primery. Metoda je tvořena opakováním těchto kroků: denaturace analyzované templátové DNA nasedání dvojice primerů annealing prodlužováním fragmentů DNA. Zmnožení fragmentu DNA je prováděno v termocykleru. tnrtb.wordpress.com
30
31 objev DNA polymerázy a první pokusy s umělou syntézou většího množství DNA (amplifikace DNA) PCR replikace in vitro PCR Polymerase Chain Reaction Polymerázová řetězová reakce 1983 Karry Mullis pracoval na projektu geneticky podmíněných chorob člověka v kalifornské firmě Cetus Corporation. Hledal novou metodu analýzy mutací DNA. Modifikoval Sangerovu metodu sekvenace DNA přidal zpětný primer pro syntézu krátkých úseků DNA. Uvědomil si, že opakováním amplifikace DNA pomocí DNA polymerázy může vést k řetězovému množení specifického segmentu genomu - PCR. Pomocí PCR můžete z jedné molekuly DNA vytvořit 100 biliónů kopií za jedno odpoledne. Potřebujete jenom zkumavku, několik jednoduchých reagencií a zdroj tepla 1984 rozpracování metody s matematikem Fredem Faloonou, patentování (Mullis et al. 1986; Mullis a Faloona 1987) 1993 Nobelova cena za chemii V devadesátých letech Cetus Corp. prodává patenty PCR a Taq firmě Hoffman La Roche za 330 miliónů $

32 Polymerázová řetězová reakce denaturace annealing prodlužování cit.vfu.cz
33 Zvyšování počtu kopií fragmentu DNA při PCR


34 PCR replikace in vitro DNA
35 PCR Polymerase Chain Reaction další rozvoj metody s použitím termostabilní polymerázy z termofilní bakterie Thermus aquaticus = zkratka Taq Pol" nebo "Taq")
36 Jak lze zjistit výsledek amplifikace Výsledek amplifikace se stanovuje gelovou elektroforézou. Elektroforeogram amplifikovaných fragmentů DNA, které umožní identifikovat odrůdy brambor.
37 PCR hlavní faktory ovlivňující průběh reakce Koncentrace dntps - stock solution 100mM - ředění na 20 a 200 mol (pro každý dntp) vyšší specifičnost a nižší frekvence zařazení nesprávného nukleotidu Koncentrace MgCl2 - optimum je 1,5-2mM, - nad 6mM klesá aktivita Taq Polymerázy - nízká koncentrace nízké výtěžky PCR - příliš vysoké koncentrace rozmazané proužky na gelu nebo zmnožení nespecifických produktů Teplota annealingu nasednutí primerů - vyšší teplota = vyšší specifičnost ke komplementární sekvenci = primer nasedá na komplementární místo s vyšší přesností = přesnější výsledek PCR - nižší teplota vhodné pro studie, kdy se používají primery odvozené od příbuzných druhů; nebezpečí vzniku nespecifických produktů (jiné úseky DNA, než které jsme chtěli získat) - zvýšení specifičnosti např. tzv. Touch down PCR v prvních cyklech vyšší teplota = specifické produkty, ale v s nižším výtěžkem = v dalších cyklech vyšší teplota annealingu = zvýší se výtěžek PCR - Koncentrace templátové DNA příliš vysoká konc. DNA může bránit PCR, zároveň se naředí inhibitory PCR ve směsi (např. chemikálie použité při izolaci)
38 PCR hlavní faktory ovlivňující průběh reakce Volba polymerázy odlišné výsledky PCR od Taq P. od různých firem - např. pro sekvenace proofreading polymerase nižší procento vložení chybného nukleotidu Aditiva - pro zvýšení efektivity PCR, stabilizaci enzymu - v pufru dodávaném s polymerázou: - detergenty (TRITON X-100, TWEEN, NP-40) - zpravidla 2 ze tří uvedených - dimetylsulfoxin nebo glycerol lepší průběh denaturace a chlazení - BSA (bovinní sérový albumin) nebo želatina - Finální koncentrace aditiv se pohybuje mezi 0,1 a 1,0% celkové reakční směsi, avšak konkrétní koncentrace je většinou nutno empiricky vyzkoušet. Kontaminace nebezpečí zejména u málo koncentrovaných nebo problematických templátů - sterilní špičky, vysoce čistá voda pro PCR, pufry, PCR zkumavky, rukavice (nikdy nesterilizovat dntps, primery, polymerázu ) - příprava PCR směsí ve flow boxu v oddělené místnosti od prostor, kde se pracuje s produkty - při přecházení z místností měnit rukavice..

39 Optimalizace reakčních podmínek PCR - Změny v protokolu vedoucí k odstranění nežádoucích nespecificit a ke zvýšení množství PCR produktu. - počet cyklů - annealingová teplota - koncentrace Mg 2+, polymerázy, templátové DNA - aditiv (např. DMSO, formamid) Marker 1 - produkt 240bp Příklad optimalizace PCR Podmínky podle literatury B PCR po optimalizaci Ta = 62 C, 30 cyklů 20uM primery Ta = 67 C, 25 cyklů 10uM primery B Marker 2 - produkt 460bp Ta = 66 C, 30 cyklů 20uM primery Ta = 66 C, 25 cyklů 10uM primery
40 RAPD - Random Amplified Polymorphic DNA Variabilita délek náhodně amplifikované DNA
41 RAPD Random Amplified Polymorphic DNA Variabilita délek náhodně amplifikované DNA PCR s jedním primerem Primer dosedne na různých místech a v různých směrech amplifikuje DNA fragmenty Identifikace např. kmenů bakterií popř. ověřování homogenity linií
42 RAPD Výhody - Velmi levné - Rychlé a snadné = není třeba znát sekvenci primeru, stačí velmi málo DNA, poměrně dost fragmentů Nevýhody převažují: - Omezena hodnota informace (dominantní marker) - Nereproducibilní DNES TREND NEUŽÍVAT RAPD MARKERY
43 AFLP - Amplified Fragment Length Polymorphism Polymorfismus délky amplifikovaných fragmentů
44 AFLP Princip metoda založená na restrikci DNA dvěma enzymy selektivní namnožení jen některých proužků vizualizace proužků na gelu Postup relativně složitý (4 fáze) 1. RESTRIKCE specifické rozštěpení DNA restrikčními endonukleázami MseI - rozpoznává 4bp dlouhou sekvenci (TTAA) EcoRI - rozpoznává 6bp dlouhou sekvenci (GAATTC) (Variabilita je dána mutací v restrikčním místě insercí - delecí mezi restrikčními místy) získáme množství fragmentů s lepivými konci = sticky ends
45 AFLP 2. ligace pomocí T4 ligázy k fragmentům přidány adaptory = oligonukleotid, jeho sekvenci známe známe sekvence konců všech fragmentů můžeme je amplifikovat PCR
46 AFLP 3. preselektivní amplifikace klasická PCR s primery komplementárními k sekvenci adaptorů, navíc 1 nukleotid směrem dovnitř amplifikovaného úseku = selekce jen ca. 1/4 fragmentů (fragmenty komplementární k primerům + fragmenty EcoRI- MseI)
47 AFLP 3. preselektivní amplifikace restrikce + ligace
48 AFLP 3. preselektivní amplifikace restrikce + ligace preamplifikace
49 AFLP 4. selektivní amplifikace -fragmentů je stále mnoho pro objektivní vyhodnocení -další redukce jejich počtu použitím primerů se 3mi selektivní nukelotidy (s přesahem "dovnitř" amplifikovaných fragmentů) - tento krok se provádí opakovaně s různými primerovými kombinacemi (primery se liší zařazenými nukleotidy na koncích -redukce na 1/256 všech fragmentů (1/16 x 1/16)
50 AFLP 5. vyhodnocení -primery jsou fluorescenčně značeny sekvenátor - Primery bez značení PAGE elektroforéza
 - přidat k roztoku z předchozího kroku -")

51 AFLP realizace AFLP - realizace 1: Restrikce - příprava restrikčního mixu (EcoI, MseI, buffer, voda) - přidat k DNA - inkubovat 2 hod při 37 C 2: Ligace - příprava ligačního mixu (LIGAZA, BUFF, ADAPTORY, VODA) - přidat k roztoku z předchozího kroku - inkubovat 2,5 hod při 37 C 3: Preamplifikace - klasická PCR - ligační roztok jako matrice Kroky 1-3 se provedou pouze 1x, dál pracujeme s preamplifikátem 4: Amplifikace - klasická PCR - matrice = preamplifikát - měníme kombinace primerů = detekujeme jiné fragmenty
52 MIKROSATELITY
53 MIKROSATELITY krátké tandemové repetice, STR (short tandem repeats) repetice jednoduchých sekvencí, SSR (simple sequence repeats) krátké, tandemově se opakující jednoduché sekvenční motivy zpravidla o délce 2-6bp prokaryota a eukaryota kódující i nekódující oblasti Vysoká mutační rychlost (u savců 10-3 až 10-4/lokus/generaci) hlavní zdroj vysoké proměnlivosti - sklouznutí nukleotidového řetězce během replikace (replication slippage). GTTCTGTCATATATATATATAT CGTACTT GTTCTGTCATATATATATATATATATATCGTACTT Alely se liší svou délkou Snadná separace pomocí elektroforézy
54 MIKROSATELITY Dokonalé = perfect repeats (jeden nepřerušený motiv): Jednonukleotidové: AAAAAAAAAAAAAAAAAA Dinukleotidové: CACACACACACACACACACA = (CA)n Trinukleotidové: CGTCGTCGTCGTCGTCGTCGT = (CGT)n Tetranukleotidové: CAGACAGACAGACAGACAGA =(CAGA)n Pentanukleotidové: AAATTAAATTAAATTAAATT =(AAATT)n Hexanukleotidové: CTTTAACTTTAACTTTAACTTTAA =(CTTTAA)n Pr: (AG)32 ; (TAT)25 ; (CAA)7 Cicer Nedokonalé = imperfect repeats (motiv je přerusen jedním nebo několika bazemi): (TC)6A(TC)13 ; (AG)12GG(AG)3 Cicer Složené = compound: (směs dokonalých nebo nedokonalých vzorů několika motivů) (AT)6(GT)42AT(GT)5(GT)10 (AT)14(AG)8 (GAA)21 (TA)23 Nejčastější motivy: mono a dinukleotidové; 3, 4 a 5ti méně
55 MIKROSATELITY GTTCTGTCATATATATATATCGTACTT celková DNA PCR specifický pár primerů
56 ELFO separace
57 MIKROSATELITY Výhody - vysoká variabilita - velká početnost a rozmístění po celém genomu - jednoduchost analýzy (mikrosatelity lze poměrně snadno studovat pomocí PCR) - robustní a reproducibilní markery - KODOMINANTNÍ MARKER, IDENTIFIKACE ALEL Nevýhody - Musíme znát sekvence primerů - Pokud neznáme, tak testovat SSR z příbuzných druhů - Optimalicace PCR - Popřípadě vyvíjet nové: EST-SSR
58 MIKROSATELITY - využití jaderné mikrosatelity (SSRs) nejlepší marker pro zhodnocení variability na populační úrovni jsou potřeba druhově specifické primery, tj. metoda použitelná pouze v případě, že pro studovaný druh byly primery již publikovány chloroplastové mikrosatelity (cpdna SSRs) vhodné pro hodnocení variability na úrovni příbuzných druhů, někdy i na vnitrodruhové úrovni dostupné jsou univerzální primery Obecně: Forenzní genetika: kriminalistika, identifikace jedinců, příbuzenské vztahy analýza rodičovství (parentage analysis) určení rodičů semen (semenáčků) v populacích identifikace klonů populačně-genetické studie genový tok, migrace historie populací
59 MIKROSATELITY - využití
60 MIKROSATELITY - využití
61 SEKVENOVÁNÍ DNA
62 SEKVENOVÁNÍ DNA - princip zjištění pořadí nukleotidů v řetězci DNA ATATATAGGCAAGGAATCTCTATTATTAAATCATT Zjištění rozdílů mezi jedincemi, geny Základní informace pro konstrukci primerů využití informace ke zjištění průběhu a rychlosti evoluce zjištění podobnosti a příbuznosti taxonů
63 SEKVENOVÁNÍ DNA - princip 1) Předstupeň: PCR s použitím dvojice primerů namnožení studovaného úseku DNA 2) sekvenační reakce použití pouze jednoho primeru kromě dntp jsou ve směsi přítomny i ddntp produkce fragmentů lišících se přesně o 1 bázi Elektroforetická separace fragmentů na gelu automatický sekvenátor
64 SEKVENOVÁNÍ DNA Aplikace - Aplikační možnosti: 1. evoluce genů (vznik alel, lokusů,redukce polymorfismu v důsledku selekce atd.) 2. vnitrodruhové (populační) studie (geografická proměnlivost, tok genů, hybridizace, fylogeografie (př. mitochondriální geny, Y chromozom) 3. mezidruhové studie (studium speciace, biogeografie) - volba správné sekvence 4. Detekce SNP = identifikace jedinců, taxonů, populací sekvenace pomáhá nalézt drobné populačně- i taxonově-specifické rozdíly Výhoda - Možnost srovnání sekvenčních dat o vašem organismu s údaji v internetových databázích Př:
65 SEKVENOVÁNÍ DNA 1977: dvě metody zjištění pořadí nukleotidů v DNA Maxamova-Gilbertova (chemická) metoda je založena na bázově-specifické chemické modifikaci a následném štěpení fragmentů DNA - dnes se neužívá Krátká sekvencedna (ds nebo ss) * na 5' konci radioaktivně označena 32 P. * vzorek se rozdělí na pět částí a) Přídavek chemikálí modifikují vždy jeden nebo dva typy nukleotidů: 1. dimethylsulfát: G 2. kys. mravenčí: A+G 3. hydrazin: C + T 4. 1,5M NaCl: C b) Přídavkem piperidinu: štěpení DNA v místě modifikace c) Elfo separace Výsledek ELFO: sekvence DNA: 5 -GTCTGCA-3 (čte se zespodu) - narazila na technické problémy s vývojem standartních molekulárních kitů - technicky příliš komplexní a příliš mnoho toxických látek
66 SEKVENOVÁNÍ DNA Sangerova metoda terminace řetězce Sangerova metoda založena na terminaci replikace nového řetězce podle matrice zkoumané sekvence dideoxynukleozidtrifosfátem (ddntp) na 3 -uhlíku deoxyribózy chybí OH-skupina a proto k nim DNA polymeráza nemůže navázat další nukleotid pokud během replikace dojde k náhodné inkorporaci dideoxynukelotidu (dda, ddc, ddg, ddt), replikace se zde zastaví
67 SEKVENOVÁNÍ DNA Sangerova metoda založená na terminace řetězce Sangerova (enzymatická) metoda Vzorek DNA se rozdělí do čtyř zkumavek a do každé se přidá specifický primer, směs všech čtyř standartních nukleotidů a DNA polymeráza Do každého ze vzorků je přidán vždy jen jeden značený dideoxynukleotid - v určitém poměru s deoxinukleotidy Př: dda + da, dg, dc, dt ddt + da, dg, dc, dt. Po ukončení amplifikace separace pomocí denaturující PAGE v dlouhém (sekvenačním) gelu + detekce autoradiograficky výsledné pořadí nukleotidů se odečte
68 SEKVENOVÁNÍ DNA - zefektivnění Sangerovy metody vývoj kapilárních sekvenátorů Syntéza DNA je v principu totožná s asymetrickou PCR (tj. s jedním primerem) v normálním termocykleru, s extenzí řetězce Taq polymerázou. k detekci produktů jsou užívány fluorescenčně značené ddntp = každý dideoxynukleotid má jinou značku tzv. Dye-terminator sequencing Př. ABI PRISM Big Dye Terminator v 3.1. Ready Reaction Cycle Sequencing Kit Polymerizační reakce probíhá v jednom vzorku během PCR dochází k produkci fragmentů lišících se o jednu bázi detekce produktů probíhá během elektroforetické migrace v kapiláře pomocí laserového detektoru sekvence je zaznamenána přímo do paměti počítače bp/run

69 SEKVENOVÁNÍ DNA - zefektivnění Sangerovy metody vývoj kapilárních sekvenátorů Vzorek s fluorescenčně značenými fragmenty po PCR
70 SEKVENOVÁNÍ DNA
71 SEKVENOVÁNÍ DNA - zefektivnění metody vývoj kapilárních sekvenátorů Kapiláry - vnitřní průměr 50μm - naplněny gelem s nízkou viskozitou, který nahradil polyakrylamid (dříve kapiláry nešlo opakovaně používat) - kratší časy analýz - plně automatizované systémy
72
73 Automatické sekvenátory
74 Nature Reviews Microbiology 7,
75 NGS = Next Generation Sequencing Celogenomové sekvenování - paralelně probíhající sekvenační reakce - produkují tisíce miliony sekvencí v jednom běhu - čtení kratších úseků (30-300bp) - Obecný postup 1. Izolace DNA 2. Příprava DNA knihovny PCR, klonování 3. Vlastní sekvenace 4. Vyhodnocení surových dat 5. Sestavení konsenzuální sekvence
76
77 Nature Reviews Microbiology 7,
78 454 - Pyrosekvenace - první nová sekvenační metoda (1998) - princip: detekce aktivity DNA-polymerázy během syntézy DNA - Výhody: - délka čtených úseků 700bp - rychlost sekv. cyklus 10 hodin - sekvenace PCR produktů - Nevýhody - překryvy homopolymerních oblastí - cena
79 454 sekvenování úseky 300 bp - čteno 400 tisíc fragmentů najednou Fragmentace DNA bp Ligace adaptorů - B adaptor má biotinovou značku pro přichycení na streptavidinem obalenou magnetickou kuličku Pico-titer plate (sekvenační čip - obsahuje 1,6 mil jamek širokých 44μm - průměr DNA kuličky je 26 μm -přídavek menších kuliček dvou typů jeden obsahuje chemikálie pro pyrosekvenaci, druhý typ fixuje kuličky s DNA Emulzní PCR - DNA kuličky do roztoku oleje a vody - Protřepání = vznik mikroreaktoru okolo kuličky Sekvenační reakce - Na destičku se cyklicky nanáší roztok polymerázy a vždy jednoho nukleotidu viz. pyrosekvenace Záznam a zpracování dat
80 Pyrosekvenování 1. začlenění nukleotidu (konkrétní typ, ne směs) 2. uvolnění pyrofosfátu (PPi) 3. Vznik ATP Enzym ATP sulfuryláza katalyzuje vznik ATP reakcí PPi s adenosinfosfosulfátem (APS) 4. Emise záření: enzym luciferáza spotřebuje vzniklý ATP na oxidaci luciferinu záblesk 5. Detekce záblesku 6. Enzym apyráza odstraní nespotřebované nukleotidy a ATP 7. Promytí systému 8. Změna typu nukleotidu ve směsi krok č 1.

81 pico-titer plate from Roche's FLX
82 Roche 454 sequencing
83 Illumina Genome Analyzer -metoda sekvenace syntézou (SBS-sequencing by synthesis) - r. 2006; úseky 35-50bp - 1) Příprava knihovny - fragmentace - na konce dva adaptéry - ELFO: vyříznou se fragmenty bp - přečištění - 2) Amplifikace fragmentů (bridge amplification) - na sklíčku jsou oligonukleotidy komplementární k oběma adaptorům - jednovláknové fragmenty se navážou na sklíčko vytvoří můstek - dosyntetizování komplementárního vlákna - denaturace dva identické fragmenty navázané na sklíčko blízko sebe - opakování = vznik tisíců kopií těsně vedle sebe v tzv. clusterech
84 Illumina Genome Analyzer - 3) sekvenační reakce - v každém cyklu NARÁZ všechny nukleotidy - mají fluorescenční značku a modifikovaný 3 konec (brání zařazení více nukleotidů v jednom kroku) - zařazení nukleotidu provázené emisí světla, charakteristického pro daný nukleotid - detekce - odstranění fluoresc. značky a odblokování 3 konce - opakování postupu - x cyklů - zpracování dat
85
86 Illumina sequencing
87
 + malé množství vzorku - čtené úseky: 100 bp - náročnost zpracování dat a rychlost analýzy")
88 SOLiD sequencing Sekvenace pomocí LIGACE krátkých oligo sond, které jsou fluorescenčně značeny + cena + vysoká přesnost metody (99,95%) + malé množství vzorku - čtené úseky: 100 bp - náročnost zpracování dat a rychlost analýzy (týden)
89 SOLiD sequencing 1. Příprava DNA knihovny Štěpení templátové DNA na kratší fragmenty Připojení adaptorů P1 a P2 empcr amplifikace fragmentů na magn. kuličkách modifikace 5 konce připojení na sekvenační destičku 2. Vlastní sekvenace
90 SOLiD sequencing - Sekvenační reakce Navázání univ. primeru, ligace sondy Připojení univ. primeru k adaptoru P1 Přidání oktamerních sond s fluoro. značkami 4 barvy, 16 kombinací koncových nukleotidů na 1. a 2. pozici Pokud je sonda na 1. a 2. pozici komplementární k 1. a 2. pozici templátu prodloužení řetězce DNA ligázou 2. Emise a detekce fluoresc. signálu 3. Odštěpení fluoresc. značky (štěpení sondy mezi 5. a 6. nukeotidem) 4. Opakování kroků x-krát po konec sekvence 5. Reset primeru odštěpení nově nasynt. Vlákna 6. Napojení nového univ. primeru k adaptoru P1 - primer je o jeden nukleotid kratší 7. Opakování kroku s primery (n-2) až (n-6) 8. Vyhodnocení dat dešifrování barevného kódu

91 8. Vyhodnocení dat dešifrování barevného kódu - každá báze je určena na základě dvou nezávislých sekvenačních cyklů
92
93 SOLiD sequencing
94
95
96
97
98
99
100 Odvozené metody využívající NGS sekvenace při hledání SNP polymorfismů

101 RRL sequencing Trends in Genetics April 2013, Vol. 29, No. 4
 anotace k referenčnímu genomu b) pooling vzorků a hledání SNP lokusů (putative RADtaq locus) - Kombinace snížení komplexnosti genomu restrikčním štěpením s NGS")
102 RAD-Taq sequencing - restrikce: DNA je naštípána restriktázami, - ligace: na vzniklé fragmenty je navázán adaptor, - vysokokapacitní NGS sekvenování (tagged fragment amplification) - analýza dat. - Hledání SNP a) anotace k referenčnímu genomu b) pooling vzorků a hledání SNP lokusů (putative RADtaq locus) - Kombinace snížení komplexnosti genomu restrikčním štěpením s NGS sekvenováním.
103 That s all folks!
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Analýza DNA. Co zjišťujeme u DNA DNA. PCR polymerase chain reaction. Princip PCR PRINCIP METODY PCR
 o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
Co zjišťujeme u DNA ACGGTCGACTGCGATGAACTCCC ACGGTCGACTGCGATCAACTCCC ACGGTCGACTGCGATTTGAACTCCC
 Analýza DNA Co zjišťujeme u DNA genetickou podstatu konkrétních proteinů mutace bodové, sekvenční delece/inzerce nukleotidů, chromosomové aberace (numerické, strukturální) polymorfismy konkrétní mutace,
Analýza DNA Co zjišťujeme u DNA genetickou podstatu konkrétních proteinů mutace bodové, sekvenční delece/inzerce nukleotidů, chromosomové aberace (numerické, strukturální) polymorfismy konkrétní mutace,
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární markery II. mikrosatelity, sekvenace, NGS, metody odvozené od NGS
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární markery II. mikrosatelity, sekvenace, NGS, metody odvozené od NGS
Analýza DNA. Co zjišťujeme u DNA
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Bi5130 Základy práce s lidskou adna
 Bi5130 Základy práce s lidskou adna Mgr. et Mgr. Kristýna Brzobohatá pizova@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu Bi5130 Základy práce s lidskou adna PCR polymerase
Bi5130 Základy práce s lidskou adna Mgr. et Mgr. Kristýna Brzobohatá pizova@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu Bi5130 Základy práce s lidskou adna PCR polymerase
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Mendelova genetika v příkladech. Genetické markery
 Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Genetické markery, markery DNA
 Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
Co zjišťujeme u DNA ACGGTCGACTGCGATGAACTCCC ACGGTCGACTGCGATCAACTCCC ACGGTCGACTGCGATTTGAACTCCC
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Genetické markery - princip a využití
 Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genotypování: Využití ve šlechtění a určení identity odrůd
 Molekulární přístupy ve šlechtění rostlin Aplikovaná genomika Genotypování: Využití ve šlechtění a určení identity odrůd Miroslav Valárik 14.2. 2017 Šlěchtění rostlin: Cílený výběr a manipulace s genomy
Molekulární přístupy ve šlechtění rostlin Aplikovaná genomika Genotypování: Využití ve šlechtění a určení identity odrůd Miroslav Valárik 14.2. 2017 Šlěchtění rostlin: Cílený výběr a manipulace s genomy
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Sekvenování příští generace (Next Generation Sequencing, NGS)
") Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR)
") POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
Využití molekulárních markerů v systematice a populační biologii rostlin. 10. Další metody
 Využití molekulárních markerů v systematice a populační biologii rostlin 10. Další metody Další molekulární markery trflp ISSRs (retro)transpozony kombinace a modifikace různých metod real-time PCR trflp
Využití molekulárních markerů v systematice a populační biologii rostlin 10. Další metody Další molekulární markery trflp ISSRs (retro)transpozony kombinace a modifikace různých metod real-time PCR trflp
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII. Martina Nováková, VŠCHT Praha
 MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Polymerázová řetězová reakce. Základní technika molekulární diagnostiky.
 Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION
 USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek
 Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek") Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Výzkumné centrum genomiky a proteomiky. Ústav experimentální medicíny AV ČR, v.v.i.
 Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
MOLEKULÁRNÍ TAXONOMIE - 4
 MOLEKULÁRNÍ TAXONOMIE - 4 V této přednášce si představíme metody, které získávají molekulární znaky bez použití sekvenace. Všechny tyto metody je teoreticky možné sekvenací nahradit. Oproti sekvenaci celých
MOLEKULÁRNÍ TAXONOMIE - 4 V této přednášce si představíme metody, které získávají molekulární znaky bez použití sekvenace. Všechny tyto metody je teoreticky možné sekvenací nahradit. Oproti sekvenaci celých
Amplifikační metody umožňují detekovat. k dispozici minimálně kopií DNA,
 Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
velké fragmenty střední fragmenty malé fragmenty
 velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
Sekvenování DNA. stanovení pořadí nukleotidů v molekule DNA (primární struktury)
") Sekvenování DNA stanovení pořadí nukleotidů v molekule DNA (primární struktury) Sekvencování / Sekvenování?? Sequencing / - die Sequenzierung / - Klasické techniky sekvenování 2 metody: Chemická (Maxamova-Gilbertova)
Sekvenování DNA stanovení pořadí nukleotidů v molekule DNA (primární struktury) Sekvencování / Sekvenování?? Sequencing / - die Sequenzierung / - Klasické techniky sekvenování 2 metody: Chemická (Maxamova-Gilbertova)
Molekulární genetika
 Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Molekulární genetika II zimní semestr 4. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Ivo Papoušek. Biologie 8, 2015/16
 Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Fingerprinting mikrobiálního společenstva (DGGE/TGGE, RFLP,T-RFLP, AFLA, ARDRA, (A)RISA)
RISA)") EKO/MEM - Molekulární ekologie mikroorganizmů Fingerprinting mikrobiálního společenstva (DGGE/TGGE, RFLP,T-RFLP, AFLA, ARDRA, (A)RISA) EKO/MEM - Molekulární ekologie mikroorganizmů DNA fingerprinting genetická
EKO/MEM - Molekulární ekologie mikroorganizmů Fingerprinting mikrobiálního společenstva (DGGE/TGGE, RFLP,T-RFLP, AFLA, ARDRA, (A)RISA) EKO/MEM - Molekulární ekologie mikroorganizmů DNA fingerprinting genetická
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Polymerázová řetězová reakce
 Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
PŘEHLED SEKVENAČNÍCH METOD
 PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Elektroforéza Sekvenování
 Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie.
 Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie. Připravila L.Fajkusová Online Mendelian Inheritance in Man: #229300 FRIEDREICH ATAXIA 1; FRDA *606829 FRDA GENE; FRDA Popis onemocnění
Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie. Připravila L.Fajkusová Online Mendelian Inheritance in Man: #229300 FRIEDREICH ATAXIA 1; FRDA *606829 FRDA GENE; FRDA Popis onemocnění
Izolace RNA. doc. RNDr. Jan Vondráček, PhD..
 Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.
 Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016 Molekulární markery využívané při studiu variability
Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016 Molekulární markery využívané při studiu variability
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014. Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU
 V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Polymorfizmy detekované. polymorfizmů (Single Nucleotide
 Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Molekulární metody ve studiích kořenových systémů. Jiří Košnar, 2016
 Molekulární metody ve studiích kořenových systémů Jiří Košnar, 2016 Úvod Řešení otázek: identifikace (barcoding) a kvantifikace rostlinných druhů ve společenstvu identifikace (barcoding) a kvantifikace
Molekulární metody ve studiích kořenových systémů Jiří Košnar, 2016 Úvod Řešení otázek: identifikace (barcoding) a kvantifikace rostlinných druhů ve společenstvu identifikace (barcoding) a kvantifikace
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Molekulárně biologické metody princip, popis, výstupy
 & Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
& Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
Genetické metody v zoologii
 Genetické metody v zoologii M. Macholán J. Bryja M. Vyskočilová Sylabus 1. Analýza fenotypu (signální fenotypy, epigenetické znaky, kvantitativní znaky, analýza landmarků) MM 2. Cytogenetika (analýza karyotypu,
Genetické metody v zoologii M. Macholán J. Bryja M. Vyskočilová Sylabus 1. Analýza fenotypu (signální fenotypy, epigenetické znaky, kvantitativní znaky, analýza landmarků) MM 2. Cytogenetika (analýza karyotypu,
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské. doc. RNDr. Ivan Mazura, CSc.
 Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) SNPs Odvozování a genotyping Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s problematikou hledání
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) SNPs Odvozování a genotyping Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s problematikou hledání
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
variabilita genomu bottleneck Nature Science
 variabilita genomu Nature Science genetická diverzita člověka na úrovni SNP je nízká: asi 0.1% cca. 90% variace je uvnitř populací cca. 10% mezi populacemi (kontinenty) bottleneck personal genomes James
variabilita genomu Nature Science genetická diverzita člověka na úrovni SNP je nízká: asi 0.1% cca. 90% variace je uvnitř populací cca. 10% mezi populacemi (kontinenty) bottleneck personal genomes James
ISSR (Inter simple sequence repeat) Jiří Košnar
 Jiří Košnar") ISSR (Inter simple sequence repeat) Jiří Košnar Přírodovědecká fakulta Jihočeské univerzity v Č. Budějovicích HISTORIE relativně nová metoda: Zietkiewicz E., Rafalski A., Labuda D. (1994): Genome fingerprinting
ISSR (Inter simple sequence repeat) Jiří Košnar Přírodovědecká fakulta Jihočeské univerzity v Č. Budějovicích HISTORIE relativně nová metoda: Zietkiewicz E., Rafalski A., Labuda D. (1994): Genome fingerprinting
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Metody molekulární biologie v rostlinné ekologii a systematice
 Protokoly a návody pro praktikum Metody molekulární biologie v rostlinné ekologii a systematice Laboratoř molekulární biologie rostlin, PřF JCU, Branišovská 31, České Budějovice 2008, Miroslava Herbstová,
Protokoly a návody pro praktikum Metody molekulární biologie v rostlinné ekologii a systematice Laboratoř molekulární biologie rostlin, PřF JCU, Branišovská 31, České Budějovice 2008, Miroslava Herbstová,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Ivo Papoušek. Biologie 6, 2017/18
 Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Metody studia historie populací. Metody studia historie populací. 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.
 Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.") 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva
1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Determinanty lokalizace nukleosomů
 METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
MagPurix Blood DNA Extraction Kit 200
 MagPurix Blood DNA Extraction Kit 200 Kat. č. ZP02001-48 Doba zpracování: 50-60 minut pro MagPurix 12S 50-70 minut pro MagPurix 24 Použití Souprava MagPurix Blood DNA Extraction Kit 200 je určena pro izolátor
MagPurix Blood DNA Extraction Kit 200 Kat. č. ZP02001-48 Doba zpracování: 50-60 minut pro MagPurix 12S 50-70 minut pro MagPurix 24 Použití Souprava MagPurix Blood DNA Extraction Kit 200 je určena pro izolátor
Biotechnologický kurz. III. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
Enzymy v molekulární biologii, RFLP. Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek
 Enzymy v molekulární biologii, RFLP Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek Enzymy v molekulární biologii umožňují nám provádět celou řadu přesně cílených manipulací Výhody enzymů:
Enzymy v molekulární biologii, RFLP Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek Enzymy v molekulární biologii umožňují nám provádět celou řadu přesně cílených manipulací Výhody enzymů:
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů
 Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární markery I. Isozymy, PCR, RAPD, AFLP Miloslav Kitner Katedra botaniky
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární markery I. Isozymy, PCR, RAPD, AFLP Miloslav Kitner Katedra botaniky
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Využití rekombinantní DNA při studiu mikroorganismů
 Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Genotypování markerů užitkovosti a zdraví u skotu
 Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
INTRODUCING OF SNAPSHOT METHOD FOR POLYMORPHISM DETECTION ZAVEDENÍ SNAPSHOT METODIKY PRO DETEKCI POLYMORFISMŮ
 INTRODUCING OF SNAPSHOT METHOD FOR POLYMORPHISM DETECTION ZAVEDENÍ SNAPSHOT METODIKY PRO DETEKCI POLYMORFISMŮ Civáňová K., Knoll A. Ústav genetiky, Agronomická fakulta, Mendelova zemědělská a lesnická
INTRODUCING OF SNAPSHOT METHOD FOR POLYMORPHISM DETECTION ZAVEDENÍ SNAPSHOT METODIKY PRO DETEKCI POLYMORFISMŮ Civáňová K., Knoll A. Ústav genetiky, Agronomická fakulta, Mendelova zemědělská a lesnická