Enzymy v molekulární biologii, RFLP. Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek
|
|
|
- Lukáš Růžička
- před 8 lety
- Počet zobrazení:
Transkript
1 Enzymy v molekulární biologii, RFLP Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek

2 Enzymy v molekulární biologii umožňují nám provádět celou řadu přesně cílených manipulací Výhody enzymů: vysoká specifičnost reakcí možnost pracovat s malým množstvím substrátu Klasifikace enzymů, jejichž substrátem jsou NK: A. Podle typu substrátu: DNA enzymy X RNA enzymy B. Podle typu reakce: - enzymy anabolické = polymerázy (DNA/RNA polymerázy, reverzní transkriptázy ) - enzymy katabolické = nukleázy (degradace: od konců = exonukleázy, nebo uvnitř molekuly = endonukleázy) - enzymy spojující řetězce NK = ligázy - enzymy modifikující NK (metylázy, kinázy, fosfatázy)
 - enzymy spojující řetězce NK = ligázy - enzymy modifikující NK (metylázy, kinázy,")
3 Anabolické enzymy syntetizovaná molekula DNA matrice enzymová třída příklad DNA DNA-polymerázy DNA-polymeráza I, Taq polymeráza RNA zpětné (reverzní) transkriptázy zpětná transkriptáza AMV žádná terminální transferázy terminální transferáza DNA RNA-polymerázy RNA-polymeráza T3, T7, SP6 žádná poly(a) polymerázy poly(a) polymeráza RNA
 polymerázy poly(a)")
4 Katabolické enzymy rozkládaná molekula typ degradace od konců enzymová třída exonukleázy DNA RNA specifita příklad ss exonukleáza III ds Bal31 ss S1 nukleáza ds restrikční endonukleázy (EcoRI ), DNáza I uvnitř molekuly endonukleázy od konců exonukleázy fosfodiesteráza uvnitř molekuly endonukleázy ribonukleáza A

5 Enzymy modifikující nukleovou kyselinu typ enzymu katalyzovaná reakce příklad metyláza metylace Dam-metyláza Dcm-metyláza EcoRI-metyláza kináza fosforylace T4-polynukleotid kináza fosfatáza defosforylace BAP-fosfatáza CIP-fosfatáza Enzymy spojující molekuly DNA typ enzymu DNA ligáza katalyzovaná reakce ligace příklad T4-DNA-ligáza

6 Nukleázy degradují molekuly NK rozkladem fosfodiesterových vazeb často nežádoucí narušují integritu vzorků NK Jednotlivé nukleázy se liší zejména substrátovou specifitou (DNA/RNA, ss/ds) a způsobem degradace substrátu: exonukleázy postupně odstraňují koncové nukleotidy endonukleázy štěpí vnitřní fosfodiesterové vazby uvnitř molekuly NK

 či ssdna uvnitř molekuly (endonukleázová")
7 Nukleázy Dnáza I (deoxyribonukleáza I) nespecifická, štěpí ss i dsdna vytváří jednořetězcové zlomy, od kterých se DNA dále degraduje na di-, tria tetranukleotidy fosforylované na 5 konci využití: např. odbourání DNA z roztoků RNA, analýza interakcí protein-dna Nukleáza Bal31 štěpí lineární dsdna od obou konců (exonukleázová aktivita v obou směrech) či ssdna uvnitř molekuly (endonukleázová aktivita) využití cílené zkracování dsdna, odstranění restrikčních míst, tvorba delecí
 či ssdna uvnitř molekuly (endonukleázová aktivita) využití cílené zkracování")
8 Nukleázy Nukleáza S1 specifická pro ssdna odstranění jednovláknových struktur (smyček) z dsdna/rna odstranění jednořetězcových konců DNA -> vytváří tupé konce využití: např. odbourání DNA z roztoků RNA, analýza interakcí protein-dna Nukleáza ExoIII postupné odstraňování mononukleotidů z 3 konců dsdna preferovaná dsdna s tupými či 5 přečnívajícími konci inaktivní na ssdna

9 Ribonukleázy Ribonukleáza T1 specificky degraduje RNA v místech G (štěpí za guaninovým zbytkem) využití: odstranění, sekvenování, mapování, studium struktury RNA odstranění jednovláknových struktur (smyček) z dsdna/rna Ribonukleáza A specificky degraduje ssrna v místech C a U (pyrimidiny) využití: eliminace kontaminující RNA např. při izolaci plazmidové DNA
 využití: eliminace kontaminující RNA např.")
10 Restrikční endonukleázy (RE) endonukleázy izolované z bakterií spolu s metylázami tvoří analogii jednoduchého imunitního systému; původně ochrana bakterií před cizorodou DNA omezují množení bakteriofágů DNA hostitelské buňky je před účinkem vlastní endonukleázy chráněna metylací

11 Restrikční endonukleázy (RE) Význam: příprava rekombinantních molekul DNA analýza polymorfismů na úrovni jedinců/populací/druhů/ (fingerprinting) studium struktury, organizace, exprese a evoluce genomu genové inženýrství (příprava sond, mutantů, modifikace DNA) Vlastnosti: štěpí oba řetězce dsdna sekvenčně extrémně specifické, rozeznávají určitou konkrétní sekvenci (rozpoznávací místo), obvykle palindrom 3 třídy dle charakteru štěpení, velikosti molekuly enzymu a požadavcích na kofaktory třídy I a III současně modifikační a restrikční aktivita, in vitro využívána třída II jen restrikční aktivita

12 Restrikční endonukleázy třídy I a III např. EcoK, EcoB (třída I), EcoP1 (třída III) rozpoznávají/vážou se na specifické nukleotidové sekvence, ale štěpí náhodně v místech vzdálených až několik tisíc bp (třída I) nebo 5-10 bp (třída III) od místa vazby molekulová hmotnost cca modifikace (metylace) i restrikce DNA kofaktor: ATP prakticky nevyužitelné kvůli náhodnosti štěpení
 od místa vazby molekulová hmotnost cca 300 000 modifikace (metylace) i restrikce")
13 Restrikční endonukleázy třídy II vážou se na specifickou (4-8 bp) sekvenci (často palindrom) rozpoznávací (cílové, vazebné) místo následně štěpí dsdna v restrikčním místě uvnitř vazebného místa nebo v jeho bezprostředním sousedství, opět naprosto specifické -> vznikají restrikční fragmenty DNA molekulová hmotnost známo více než 3500 restrikčních endonukleáz třídy II (s různými rozpoznávacími a restrikčními místy teoreticky dokážeme rozštěpit v libovolně zvolené sekvenci) cílové místo je rotačně symetrické důležitá je orientace cílového místa
 cílové místo je rotačně symetrické důležitá je orientace")
14

15 Restrikční endonukleázy třídy II Produkty štěpení: tupé ( blunt ) konce vznikají po štěpení obou řetězců v témže místě ostré ( sticky, přečnívající, kohezní, nerovné) konce po štěpení řetězců v různých místech vzdálených obvykle 1-4 N 5 přečnívající nebo 3 přečnívající

16

17 Restrikční endonukleázy třídy II Produkty štěpení: tupé ( blunt ) konce vznikají po štěpení obou řetězců v témže místě ostré ( sticky, přečnívající, kohezní, nerovné) konce po štěpení řetězců v různých místech vzdálených obvykle 1-4 N 5 přečnívající nebo 3 přečnívající význam přečnívající komplementární konce umožňují snadné spojení fragmentů DNA

18 Příklady cílových míst
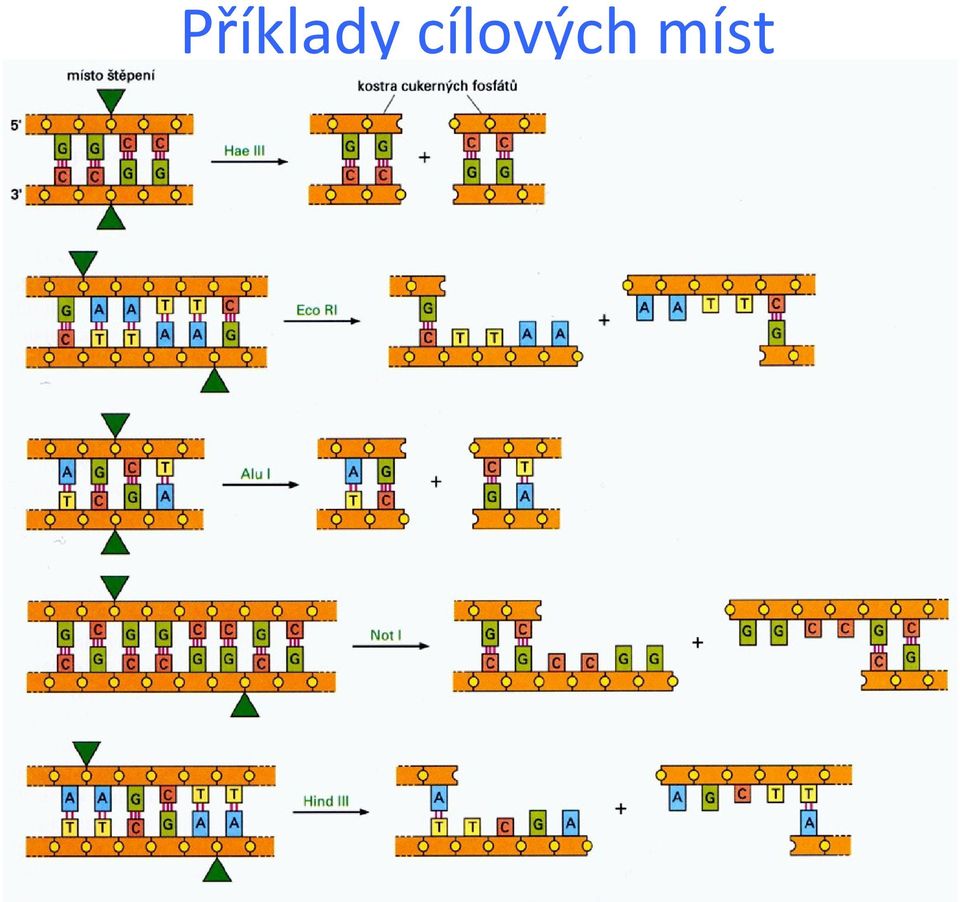
19 Restrikční endonukleázy (RE) Názvosloví: např. EcoRI 1. písmeno počáteční písmeno rodu příslušné produkující bakterie 2. a 3. písmeno první dvě písmena druhu bakterie 4. písmeno (ne vždy) označení kmene/serotypu Římská číslice pořadové číslo endonukleázy izolované z dané bakterie Izoschizomery různé RE se stejným rozpoznávacím místem, ale odlišným způsobem štěpení či jinou citlivostí k metylaci
 označení kmene/serotypu Římská číslice pořadové číslo endonukleázy izolované z")
20 Restrikční endonukleázy (RE) Relaxovaná (uvolněná) specifita: schopnost štěpit kromě cílového místa také jiné, podobné/příbuzné sekvence uplatňuje se obvykle za suboptimálních reakčních podmínek (teplota, soli ) např. EcoRI štěpí GAATTC, ale dokáže štěpit i GGATTC, GGATTT aj. Citlivost k metylaci: vyznačuje se značkami nad rozpoznávanou sekvencí + + AC štěpení je inhibováno metylací adeninu nebo cytozinu 0 0 AC štěpení není metylací adeninu nebo cytozinu ovlivněno Množství RE: konvenčně vyjadřujeme v tzv. jednotkách (unit, U) 1 U množství RE, které zcela rozštěpí 1 μg DNA fága lambda za 1 hodinu při optimální teplotě a v optimálním prostředí

21 RFLP polymorfismus délky restrikčních fragmentů Princip: DNA různých jedinců (druhů, vyšších taxonů ) se liší sekvencí liší se přítomností restrikčních míst -> restrikčními enzymy bude štěpena na různé soubory fragmentů jednotlivé soubory se můžou lišit počtem fragmentů a jejich velikostmi příklady: samec vykazuje jiný soubor fragmentů než samice, kůň jiný soubor fragmentů než kráva štěpit můžeme buď celou genomovou DNA nebo zvolený úsek (např. předem amplifikovaný pomocí PCR) variabilita spektra fragmentů záleží především na zvoleném sledovaném úseku DNA a použitém restrikčním enzymu tak nastavuji citlivost (zda rozeznám jedince, druhy )
22 RFLP polymorfismus délky restrikčních fragmentů Postup: 1) izolace genomové DNA/amplifikace zájmového úseku pomocí PCR 2) štěpení restrikční endonukleázou (případně více po sobě) 3) separace vzniklých fragmentů DNA pomocí gelové elektroforézy RFLP vzniká přestavbami sekvencí - inzercemi - delecemi - substitucemi bazí uvnitř restrikčních míst
23 RFLP polymorfismus délky restrikčních fragmentů V hygieně potravin: velmi tradiční a často využívaná metoda rozlišení např. masa pocházejícího z různých druhů zvířat, či různé druhy ryb, drůbeže, aniž by bylo nutno fragment DNA přímo sekvenovat zpravidla využívána varianta PCR-RFLP (neštěpíme celou genomovou DNA, ale jen vybraný úsek, který amplifikujeme - namnožíme) výběrem úseku určujeme, co/na jaké úrovni chceme rozlišit (druhy ryb? Druhy drůbeže? Plemena skotu?) jednoduché, snadno přístupné v jakékoli laboratoři nevýhoda: nebude dobře fungovat např. na masové směsi (vzniknou obtížně interpretovatelné směsné vzory fragmentů), je potřeba čistý vzorek
24 (a) EcoRI1 and (b) BamH1 restriction profiles obtained from PCRRFLP analysis of centrometric satellite DNA in cooked meat from nine animal species. Lane numbers: 1=molecular marker of 1000 bp; 2=taurine cattle; 3=zebu cattle; 4=banteng; 5=buffalo; 6=bison; 7= wisent; 8=water buffalo; 9=African buffalo; 10=Indian cattle.
25 Polymerázy syntéza nukleových kyselin postupným doplňováním nukleotidů k 3 -OH konci existující molekuly NA (obvykle primeru) syntéza probíhá dle matrice (templátu) ssdna nebo RNA polymeráza tak vytváří komplementární kopii templátového řetězce není známa polymeráza prodlužující 5 konec DNA
26 DNA polymeráza I izolována z Escherichia coli 3 aktivity 3 katalyzované reakce: 1) Aktivita polymerázová (5 ->3 jiná být nemůže) 2) 3 ->5 exonukleázová aktivita 3) 5 ->3 exonukleázová aktivita Polymerace: - prodlužování polynukleotidového řetězce, tvorba fosfodiesterové vazby mezi 3 OH skupinou rostoucího řetězce a 5 P dntp Exonukleázová aktivita hydrolýza DNA v obou směrech Degradace DNA ve směru 5 -> 3 a současná syntéza degradovaného řetězce (tj. kombinace 1 a 3) in vivo oprava chybně začleněných nukleotidů praktické využití při značení DNA
27 Polymerace DNA
28 Tři aktivity DNA polymerázy I
29 Klenowův fragment DNA polymerázy I vzniká odstraněním N-koncové části AK řetězce postrádá 5 ->3 exonukleázovou aktivitu komerčně využívaný: syntéza dsdna ze ss templátů, kdy nesmí docházet k odbourání primerů (např. enzymatické sekvenování) doplnění zkrácených 3 konců fragmentů DNA vytváření tupých konců vhodných pro klonování odstranění přečnívajících 3 konců tentýž účel; ovšem odstraňování od 3 konců má tendenci pokračovat; za přítomnosti nukleotidů je to vyrovnáno polymerační aktivitou -> tupé konce
30 T4 DNA polymeráza DNA polymeráza fága T4 polymerázová aktivita 5 3 exonukleázová aktivita 3 5 (200 x vyšší než u Klenowova enzymu) využití: značení 3 konců DNA: exonukleázová aktivita degraduje dvouřetězcovou DNA odbourání 3 konců obou řetězců. Po přidání značených dntp jsou konce doplněny polymerační aktivitou enzymu.
31 Termostabilní DNA polymerázy Taq DNA polymeráza izolována v roce 1976 z bakterie Thermus aquaticus horké prameny nepodléhají denaturaci teplem esenciální podmínka pro polymerázovou řetězovou reakci (PCR) běžné polymerázy by nepřečkaly denaturační krok (Taq - poločas života cca 1,6 hod při 95 C) maximum katalytické aktivity mezi 75 a 80 C, při nižších teplotách klesá další termostabilní DNA polymerázy: Pfu (Pyrococcus furiosus, nejvyšší přesnost), Vent (Thermococcus litoralis, dlouhý poločas života 7 hod v 95 C) dnes produkované geneticky upravenými bakteriemi, upraveny tak, aby měly co nejvýhodnější vlastnosti (např. hot-start aktivace vysokou teplotou, brání předčasné katalýze)
32 Reverzní (zpětná) transkriptáza RNA-dependentní DNA polymeráza tj. přepis genetické informace z RNA do DNA tak jako ostatní polymerázy vyžaduje přítomnost primeru a templátu v prvním kroku vzniká hybrid RNA/DNA, po odstranění RNA (hydroxidy, RNáza H) může katalyzovat i syntézu druhého vlákna syntéza druhého vlákna může být zajištěna i běžnou DNA polymerázou původní zdroj retroviry; běžně využívají ve svém životním cyklu Aktivity: polymerázová: u retrovirů pouze zpětný přepis RNA, v laboratoři pro transkripci ssrna i ssdna. V obou případech nutná přítomnost primeru 5 -> 3 exoribonukleázová 3 -> 5 exoribonukleázová a endoribonukleázová (RNáza H): zajišťuje degradaci RNA z hybridů RNA-DNA, které vznikají při zpětné transkripci.
33 Reverzní transkriptáza Využití: tvorba komplementární DNA (cdna) - nespecifická (primer: oligo dt) - specifická (primer: komplementární k 3 oblasti hledané mrna)
34 Syntéza cdna např. pro studium genové exprese, tj. těch genů, které jsou v dané tkáni právě aktivní
35 Další polymerázy Bakteriofágové DNA-dependentní RNA polymerázy transkripce in vitro tvorba přesně definovaných RNA, tvorba značené RNA (např. sondy pro hybridizaci) příprava templátu pro translaci in vitro studium struktury a funkce RNA Terminální transferázy také při syntéze cdna (jako reverzní transkriptáza) dokáží prodloužit 3 konec libovolnými nukleotidy (nepotřebuje matrici) využití: jeden ze způsobů začlenění cílového úseku DNA do vektoru (3 konce vektoru prodloužíme o (G)n, 3 konce fragmentu o (C)n, budou k sobě komplementární spojí se) Poly(A) polymeráza katalyzuje přidání asi 200 A na 3 konec mrna při sestřihu eukaryotické RNA, nepotřebuje matrici využití před syntézou cdna prostřednictvím primerů oligo (dt) pak lze příslušnou RNA zpětnou transkriptázou převést do cdna
36 Enzymy modifikující DNA modifikují DNA přidáním nebo odebráním určitých chemických skupin Alkalická fosfatáza odstraňuje fosfátové skupiny z 5 konců DNA - defosforylace využití fragmenty DNA zbavené koncových fosfátů nemohou být spojeny ligací brání recirkularizaci prázdného plazmidového vektoru usnadnění klonování; nebo defosforylace DNA před značením radioaktivním fosfátem defosforylace proteinů Polynukleotidová kináza T4 fosforylace 5 OH skupin polynukleotidů (ss i ds DNA i RNA) využití to značení radioaktivním fosfátem (viz výše)
37 Ligázy katalyzují spojení fragmentů DNA s: Ostrými (kohezními, nerovnými) konci (sticky ends) Tupými konci (blunt ends) nezastupitelná role in vivo při procesech replikace, rekombinace, reparace existují i RNA-ligázy in vitro široké využití, např. pro klonování DNA (první fáze tvorba rekombinantních molekul) např. T4-DNA-ligáza (ostré i tupé), bakteriální DNA ligáza (jen ostré)
38 Tvorba rekombinantní DNA in vitro
39 Ligázy tupé konce Nižší účinnost ligace, protože setkání konců DNA je náhodné -> nutná vyšší koncentrace DNA Umožňuje kombinaci fragmentů z různých restrikcí (štěpení), ale produkty jsou heterogenní ostré konce Vyšší účinnost ligace (pokud přesahy odpovídající), vznikají homogenní produkty Úprava konců fragmentů: ostré na tupé: a) doplněním chybějících nukleotidů ( filling in ) b) odstraněním přečnívající sekvence ( trimming back ) tupé na ostré: a) linkery b) adaptéry
40 Linkery a adaptéry Linkery krátké syntetické oligonukleotidy, plně komplementární samy k sobě (např. CCGGATCCGG) v ds stavu tedy mají tupé konce nesou cílovou sekvenci pro RE (např. BamHI GGATCC) použití: spojení původně nekompatibilních molekul: 1. připojení linkeru k tupým koncům DNA ligací 2. štěpení RE vytvoří se ostré konce DNA na obou spojovaných fragmentech 3. spojení teď už kompatibilních molekul DNA ligací obecně umožňují přidání žádoucích konců/sekvencí k jakékoliv molekule DNA Adaptéry dvojice krátkých syntetických oligonukleotidů, které jsou částečně komplementární párují se v krátký fragment dsdna s přečnívajícími konci pak spojení s jiným fragmentem s tupým koncem (či nevhodným ostrým) výhoda ostré konce získány bez štěpení RE (problém linkerů, pokud je restrikční místo v klonované sekvenci)
41 Linkery
42 Linkery a adaptéry Linkery krátké syntetické oligonukleotidy, plně komplementární samy k sobě (např. CCGGATCCGG) v ds stavu tedy mají tupé konce nesou cílovou sekvenci pro RE (např. BamHI GGATCC) použití: spojení původně nekompatibilních molekul: 1. připojení linkeru k tupým koncům DNA ligací 2. štěpení RE vytvoří se ostré konce DNA na obou spojovaných fragmentech 3. spojení teď už kompatibilních molekul DNA ligací obecně umožňují přidání žádoucích konců/sekvencí k jakékoliv molekule DNA Adaptéry dvojice krátkých syntetických oligonukleotidů, které jsou částečně komplementární párují se v krátký fragment dsdna s přečnívajícími konci pak spojení s jiným fragmentem s tupým koncem (či nevhodným ostrým) výhoda ostré konce získány bez štěpení RE (problém linkerů, pokud je restrikční místo v klonované sekvenci)
43
Metody molekulární biologie
 školní rok 2010/2011, kurz Bi6400 Metody molekulární biologie prof. Jan Šmarda doc. Roman Pantůček Ústav experimentální biologie Přírodovědecká fakulta MU Osnova kurzu manipulace s nukleovými kyselinami:
školní rok 2010/2011, kurz Bi6400 Metody molekulární biologie prof. Jan Šmarda doc. Roman Pantůček Ústav experimentální biologie Přírodovědecká fakulta MU Osnova kurzu manipulace s nukleovými kyselinami:
Enzymy používané v molekulární biologii
 Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Enzymy používané v molekulární biologii
 Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Ivo Papoušek. Biologie 8, 2015/16
 Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Využití restriktáz ke studiu genomu mikroorganismů
 Využití restriktáz ke studiu genomu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartos.milan@atlas.cz Přírodovědecká fakulta MU, 2017 Obsah přednášky 1) Definice restrikčních endonukleáz, jejich přirozená
Využití restriktáz ke studiu genomu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartos.milan@atlas.cz Přírodovědecká fakulta MU, 2017 Obsah přednášky 1) Definice restrikčních endonukleáz, jejich přirozená
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Amplifikační metody umožňují detekovat. k dispozici minimálně kopií DNA,
 Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
Polymerázová řetězová reakce. Základní technika molekulární diagnostiky.
 Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Determinanty lokalizace nukleosomů
 METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Ivo Papoušek. Biologie 6, 2017/18
 Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Klonování gen a genové inženýrství
 Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Využití rekombinantní DNA při studiu mikroorganismů
 Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
a) Primární struktura NK NUKLEOTIDY Monomerem NK jsou nukleotidy
 Primární struktura NK NUKLEOTIDY Monomerem NK jsou nukleotidy") 1 Nukleové kyseliny Nukleové kyseliny (NK) sice tvoří malé procento hmotnosti buňky ale významem v kódování genetické informace a její expresí zcela nezbytným typem biopolymeru všech živých soustav a)
1 Nukleové kyseliny Nukleové kyseliny (NK) sice tvoří malé procento hmotnosti buňky ale významem v kódování genetické informace a její expresí zcela nezbytným typem biopolymeru všech živých soustav a)
Nukleové kyseliny Replikace DNA Doc. MVDr. Eva Bártová, Ph.D.
 Nukleové kyseliny Replikace DNA 2013 Doc. MVDr. Eva Bártová, Ph.D. Nukleové kyseliny 7% cytozin Monomer: NUKLEOTID, tvoří jej: uracil kyselina fosforečná pentóza (ribóza, deoxyribóza) tymin organická dusíkatá
Nukleové kyseliny Replikace DNA 2013 Doc. MVDr. Eva Bártová, Ph.D. Nukleové kyseliny 7% cytozin Monomer: NUKLEOTID, tvoří jej: uracil kyselina fosforečná pentóza (ribóza, deoxyribóza) tymin organická dusíkatá
Izolace, klonování a analýza DNA
 Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek
 Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek") Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Polymorfizmy detekované. polymorfizmů (Single Nucleotide
 Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Molekulární biotechnologie č.9. Cílená mutageneze a proteinové inženýrství
 Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Zdrojem je mrna. mrna. zpětná transkriptáza. jednořetězcová DNA. DNA polymeráza. cdna
 Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Výuka genetiky na PřF OU K. MALACHOVÁ
 Výuka genetiky na PřF OU K. MALACHOVÁ KATEDRA BIOLOGIE A EKOLOGIE BAKALÁŘSKÉ STUDIJNÍ PROGRAMY Experimentální Systematická Aplikovaná (prezenční, kombinovaná) Jednooborová Dvouoborová KATEDRA BIOLOGIE
Výuka genetiky na PřF OU K. MALACHOVÁ KATEDRA BIOLOGIE A EKOLOGIE BAKALÁŘSKÉ STUDIJNÍ PROGRAMY Experimentální Systematická Aplikovaná (prezenční, kombinovaná) Jednooborová Dvouoborová KATEDRA BIOLOGIE
Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
 MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
Izolace RNA. doc. RNDr. Jan Vondráček, PhD..
 Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Molekulární genetika
 Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Polymerázová řetězová reakce
 Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Metody studia exprese mrna. jádro a genová exprese 2007
 Metody studia exprese mrna Buněčné jádro a genová exprese 2007 Aktivita genu je primárn ě vyjád ř ena jeho transkripcí-prvním krokem vedoucím k syntéze kódovaného proteinu. Cíle metod Ur č ení mno ž ství
Metody studia exprese mrna Buněčné jádro a genová exprese 2007 Aktivita genu je primárn ě vyjád ř ena jeho transkripcí-prvním krokem vedoucím k syntéze kódovaného proteinu. Cíle metod Ur č ení mno ž ství
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII. Martina Nováková, VŠCHT Praha
 MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
Přednáška kurzu Bi4010 Základy molekulární biologie, 2016/17 Replikace DNA
 Přednáška kurzu Bi4010 Základy molekulární biologie, 2016/17 Replikace DNA Jan Šmarda Ústav experimentální biologie, PřF MU 1 Buněčné dělení a reprodukce každá buňka potřebuje svou úplnou sadu genů: rodičovská
Přednáška kurzu Bi4010 Základy molekulární biologie, 2016/17 Replikace DNA Jan Šmarda Ústav experimentální biologie, PřF MU 1 Buněčné dělení a reprodukce každá buňka potřebuje svou úplnou sadu genů: rodičovská
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Mendelova genetika v příkladech. Genetické markery
 Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Přijímací test navazující magisterské studium Molekulární a buněčná biologie
 Přijímací test navazující magisterské studium Molekulární a buněčná biologie 14. června 2016 Číslo uchazeče: Poznámky k řešení testu: Doba řešení: 60 min Správná je jen 1 odpověď, která je hodnocena 1
Přijímací test navazující magisterské studium Molekulární a buněčná biologie 14. června 2016 Číslo uchazeče: Poznámky k řešení testu: Doba řešení: 60 min Správná je jen 1 odpověď, která je hodnocena 1
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR)
") POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA
 RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
Příprava rekombinantních molekul pro diagnostické účely
 1 Příprava rekombinantních molekul pro diagnostické účely doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 2 Obsah přednášky 1) Pojem rekombinantní DNA 2) Historické milníky
1 Příprava rekombinantních molekul pro diagnostické účely doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 2 Obsah přednášky 1) Pojem rekombinantní DNA 2) Historické milníky
Chemická reaktivita NK.
 Chemické vlastnosti, struktura a interakce nukleových kyselin Bi7015 Chemická reaktivita NK. Hydrolýza NK, redukce, oxidace, nukleofily, elektrofily, alkylační činidla. Mutageny, karcinogeny, protinádorově
Chemické vlastnosti, struktura a interakce nukleových kyselin Bi7015 Chemická reaktivita NK. Hydrolýza NK, redukce, oxidace, nukleofily, elektrofily, alkylační činidla. Mutageny, karcinogeny, protinádorově
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
v raném stádiu se embryo rozpadlo do dvou skupin buněk správná odpověď: dvojčata obsahují kopie stejných rodičovských
 Replikace DNA Jsou monozygotní dvojčata identická? vyvinula se z jednoho oplozeného vajíčka v raném stádiu se embryo rozpadlo do dvou skupin buněk obě skupiny buněk prodělaly úplný vývoj a dozrály do úplných
Replikace DNA Jsou monozygotní dvojčata identická? vyvinula se z jednoho oplozeného vajíčka v raném stádiu se embryo rozpadlo do dvou skupin buněk obě skupiny buněk prodělaly úplný vývoj a dozrály do úplných
Genetické markery, markery DNA
 Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
studium množství určitého transkriptu v daném vzorku a v množství dané molekuly mrna v dané buňce a v daném
 Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc.
 Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Molekulární genetika II zimní semestr 4. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
4) pokračování struktury nukleových kyselin
 pokračování struktury nukleových kyselin") Denaturace a renaturace DNA 4) pokračování struktury nukleových kyselin Genofor, chromozom, genom Genofor struktura nesoucí geny seřazené za sebou (DNA nebo RNA) a schopná replikace. U prokaryot, eukaryot
Denaturace a renaturace DNA 4) pokračování struktury nukleových kyselin Genofor, chromozom, genom Genofor struktura nesoucí geny seřazené za sebou (DNA nebo RNA) a schopná replikace. U prokaryot, eukaryot
Izolace nukleových kyselin
 Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
První testový úkol aminokyseliny a jejich vlastnosti
 První testový úkol aminokyseliny a jejich vlastnosti Vysvětlete co znamená pojem α-aminokyselina Jaký je rozdíl mezi D a L řadou aminokyselin Kolik je základních stavebních aminokyselin a z čeho jsou odvozeny
První testový úkol aminokyseliny a jejich vlastnosti Vysvětlete co znamená pojem α-aminokyselina Jaký je rozdíl mezi D a L řadou aminokyselin Kolik je základních stavebních aminokyselin a z čeho jsou odvozeny
Genetické markery - princip a využití
 Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
SYNTETICKÉ OLIGONUKLEOTIDY
 Oddělení funkční genomiky a proteomiky Přírodovědecká fakulta Masarykovy university SYNTETICKÉ OLIGONUKLEOTIDY Hana Konečná CENTRÁLNÍ LABORATOŘ Masarykovy Univerzity v Brně ODDĚLENÍ FUNKČNÍ GENOMIKY A
Oddělení funkční genomiky a proteomiky Přírodovědecká fakulta Masarykovy university SYNTETICKÉ OLIGONUKLEOTIDY Hana Konečná CENTRÁLNÍ LABORATOŘ Masarykovy Univerzity v Brně ODDĚLENÍ FUNKČNÍ GENOMIKY A
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Nukleové kyseliny (NK)
") Eva Roubalová B10 2007/2008 Předmět: - Obecná biologie - Biologie a genetika Zdroj velké části materiálů: učebnice Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava
Eva Roubalová B10 2007/2008 Předmět: - Obecná biologie - Biologie a genetika Zdroj velké části materiálů: učebnice Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava
Analýza DNA. Co zjišťujeme u DNA
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Analýza DNA. Co zjišťujeme u DNA DNA. PCR polymerase chain reaction. Princip PCR PRINCIP METODY PCR
 o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
Funkční specializace dnes: nukleové kyseliny uchovávají genet. informaci bílkoviny mají strukturní a katalytickou fci
 Evoluce RNA Funkční specializace dnes: nukleové kyseliny uchovávají genet. informaci bílkoviny mají strukturní a katalytickou fci Po určité období měl obě funkce jeden typ sloučenin, RNA - informační i
Evoluce RNA Funkční specializace dnes: nukleové kyseliny uchovávají genet. informaci bílkoviny mají strukturní a katalytickou fci Po určité období měl obě funkce jeden typ sloučenin, RNA - informační i
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
ší šířen METODY ANALÝZY NUKLEOVÝCH KYSELIN Polymerázová řetězová reakce
 METODY ANALÝZY NUKLEOVÝCH KYSELIN Polymerázová řetězová reakce Většina metod analýzy DNA využívá možnost amplifikace DNA v in vitro podmínkách. Polymerázová řetězová reakce - PCR (polymerase chain reaction)
METODY ANALÝZY NUKLEOVÝCH KYSELIN Polymerázová řetězová reakce Většina metod analýzy DNA využívá možnost amplifikace DNA v in vitro podmínkách. Polymerázová řetězová reakce - PCR (polymerase chain reaction)
Lékařská chemie a biochemie modelový vstupní test ke zkoušce
 Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
ZDRAVOTNÍ NEZÁVADNOST POTRAVIN
 ZDRAVOTNÍ NEZÁVADNOST POTRAVIN Možnosti stanovení Listeria monocytogenes popis metod a jejich princip Mária Strážiková Aleš Holfeld Obsah Charakteristika Listeria monocytogenes Listerióza Metody detekce
ZDRAVOTNÍ NEZÁVADNOST POTRAVIN Možnosti stanovení Listeria monocytogenes popis metod a jejich princip Mária Strážiková Aleš Holfeld Obsah Charakteristika Listeria monocytogenes Listerióza Metody detekce
Hybridizace. doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz
 Hybridizace doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Způsoby provedení hybridizace 2) Hybridizace v roztoku 3) Příprava značených sond 4) Hybridizace
Hybridizace doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Způsoby provedení hybridizace 2) Hybridizace v roztoku 3) Příprava značených sond 4) Hybridizace
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
DETEKCE A IDENTIFIKACE FYTOPATOGENNÍCH BAKTERIÍ METODOU PCR-RFLP
 DETEKCE A IDENTIFIKACE FYTOPATOGENNÍCH BAKTERIÍ METODOU PCR-RFLP Polymerázová řetězová reakce (PCR) je in vitro metoda pro enzymatickou syntézu definované sekvence DNA. Reakce využívá dvou oligonukleotidových
DETEKCE A IDENTIFIKACE FYTOPATOGENNÍCH BAKTERIÍ METODOU PCR-RFLP Polymerázová řetězová reakce (PCR) je in vitro metoda pro enzymatickou syntézu definované sekvence DNA. Reakce využívá dvou oligonukleotidových
Molekulární základ dědičnosti
 Molekulární základ dědičnosti Dědičná informace je zakódována v deoxyribonukleové kyselině, která je uložena v jádře buňky v chromozómech. Zlomovým objevem pro další rozvoj molekulární genetiky bylo odhalení
Molekulární základ dědičnosti Dědičná informace je zakódována v deoxyribonukleové kyselině, která je uložena v jádře buňky v chromozómech. Zlomovým objevem pro další rozvoj molekulární genetiky bylo odhalení
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským