Sekvenování DNA. stanovení pořadí nukleotidů v molekule DNA (primární struktury)
|
|
|
- Martina Dostálová
- před 6 lety
- Počet zobrazení:
Transkript
1 Sekvenování DNA stanovení pořadí nukleotidů v molekule DNA (primární struktury)
2 Důvody řešení genomových projektů Genomika modelových organismů (de novo sekvenování) Analýza mutací a SNP, výzkum genetických onemocnění (resekvenování genomů) Identifikace mikroorganismů (de novo sekvenování a resekvenování) Analýza transkriptomu (RNAseq) DNA metylační studie (medip-seq, bisulfite treated DNA) Studie transkripčních faktorů, proteinů vázajících se na DNA (ChIPseq) mirnas, sirna, pirna, trf, aj (small RNA seq) Metagenomika (16S rrna, celé metagenomy)
3 Techniky sekvenování Klasické techniky: Chemická (Maxamova-Gilbertova) Již se nepoužívá Enzymová (Sangerova) V současnosti se používá automatická varianta Metody sekvenování nové generace (next generation sequencing, NGS) 454, IonTorrent, Illumina, SoLiD Metody sekvenování třetí generace Helicos, PacBio, NanoPore
4 Vývoj sekvenačních metod Klasické sekvenování kapilární elektroforézou Vyžaduje velký počet kopií vstupního materiálu DNA jako templátu pro přípravu jednořeťězců Sekvenování nové generace Jako templát slouží jediná molekula, která je amplifikována pro získání dostatečného signálu, produkuje krátká čtení, možnost kvantitativní analýzy (SNP) Sekvenování třetí generace Nevyužívá amplifikace za účelem zvýšení signálu, produkuje dlouhá čtení
5 Projekt Výběr platformy Příprava knihovny Sekvenování Kontrola kvality Alignment Assembly Vizualizace a analýza Normalizace, srovnání knihoven Hledání genů, variant, vazebných míst, Diferenciální analýza, exprese, metabolomika,.
6 Základní požadavky pro úspěšné stanovení sekvence: Příprava knihovny pro sekvenování - DNA spřesně definovanými konci s místem pro vazbu primeru s koncovou značkou Příprava variant fragmentů ssdna lišících se svou délkou o jeden nukleotid Přesné rozdělení těchto fragmentů na základě jejich délky, detekce připojené koncové báze
7 Chemická metoda (Maxam-Gilbert) Sekvence je odvozena z molekuly DNA, která se chemicky degraduje v místech, kde se vyskytuje báze určitého typu na fragmenty. Ty se následně oddělují elektroforézou.
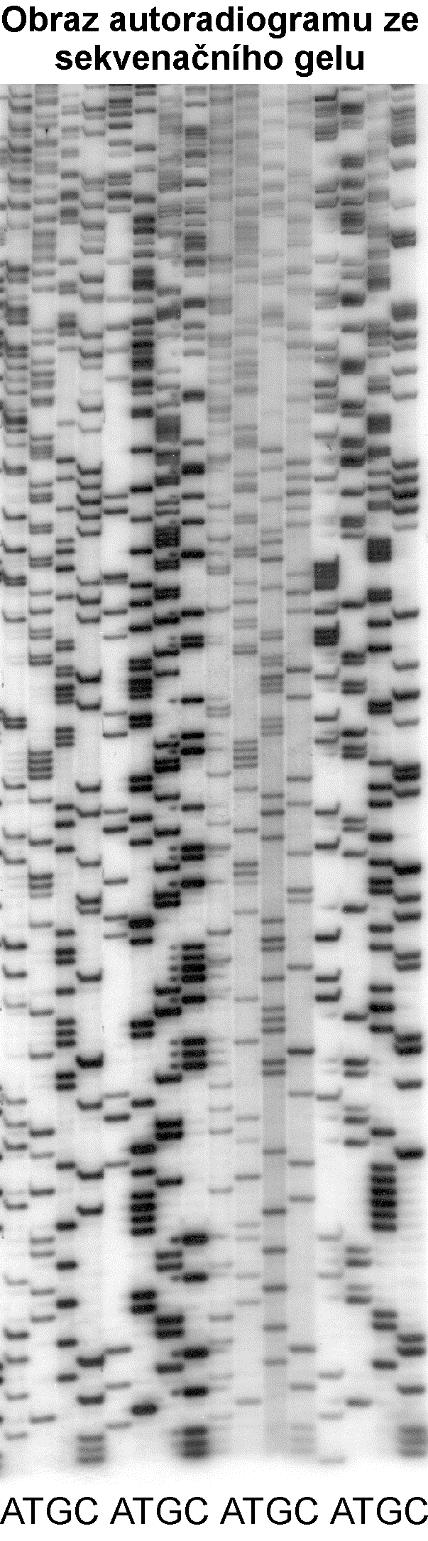
8 Chemická metoda (Maxam-Gilbert) Postup: 1. Příprava značené jednořetězcové DNA, jejíž sekvence má být stanovena 2. Rozdělení vzorku DNA do 4 částí 3. Chemická modifikace jednoho nebo dvou typů bází v náhodných místech molekuly DNA v každém vzorku 4. Štěpení molekuly DNA v místech, kde došlo k modifikaci bází 5. Elektroforéza fragmentů DNA v denaturačním polyakrylamidovém gelu 6. Autoradiografická (nebo jiná) detekce fragmentů DNA
9
10 Princip modifikace báze a štěpení
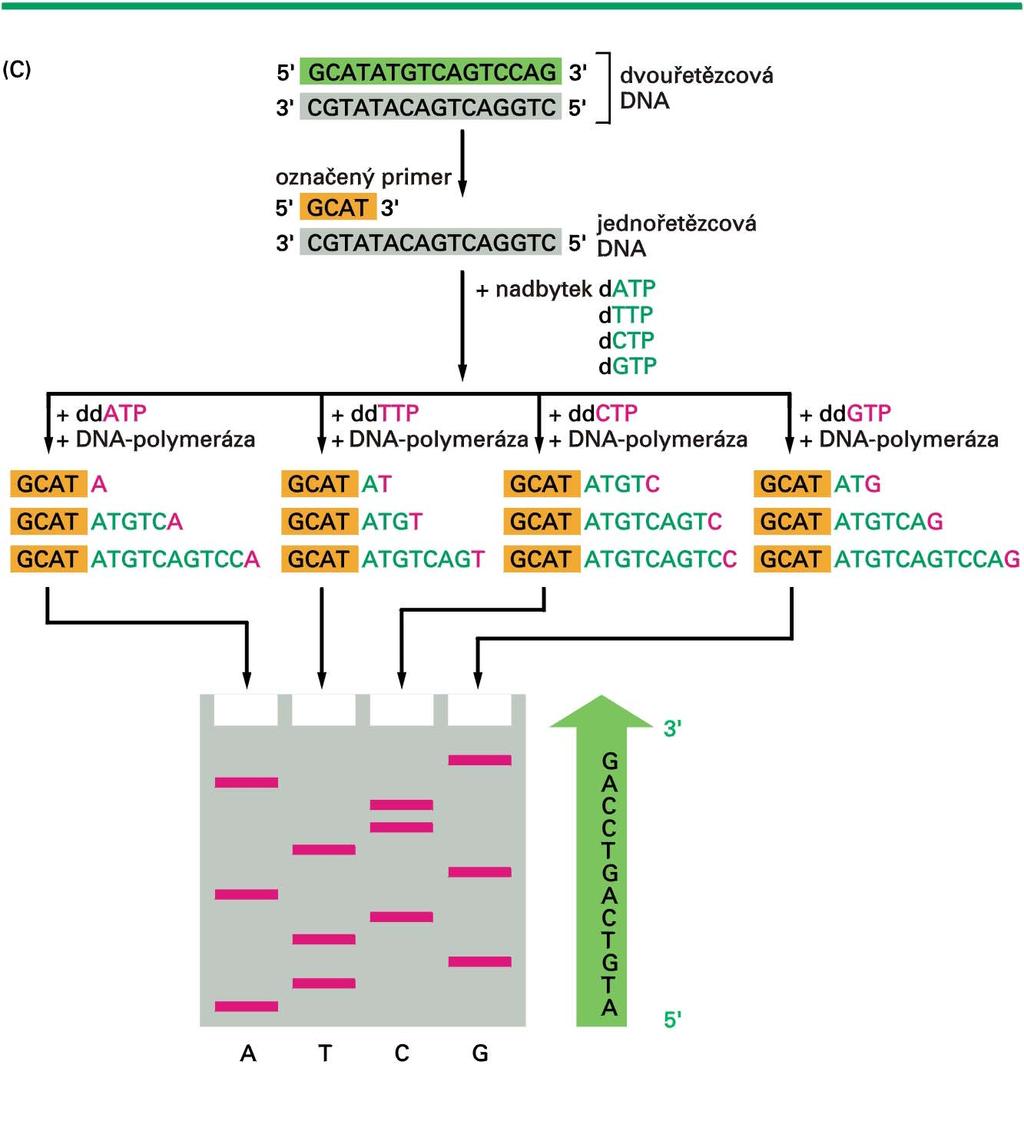
11 Postup enzymatického sekvenování DNA 1. Příprava ssdna jejíž sekvence má být stanovena 2. Rozdělení do 4 vzorků 3. Reakce s DNA polymerázou při níž se začlení do syntetizované DNA místo normálního deoxyribonukleosidtrifosfátu jeho analog, který působí jako koncový inhibitor syntézy DNA - dideoxynukleosidtrifosfát (ddntp). V každém ze 4 vzorků vzniknou fragmenty, které jsou zakončeny příslušným ddntp (ddctp, ddgtp, ddatp a ddttp). Reakce obsahuje: molekulu DNA značený primer při pojující se k části molekuly DNA (místo odkud začínáme stanovovat sekvenci) směs obsahující 4 normální nukleotidy jeden ddntp (v každém ze 4 vzorků je jiný) DNA polymerázu 4. Denaturace produktů 5. Elektroforetická separace umožňující oddělení fragmentů lišících se délkou o jednu bázi 6. Detekce fragmentů, které nesou označený konec

12 Dideoxynukleotidy nemají hydroxyl na 3 -konci
13 Pokud je dideoxynukleotid inkorporován do syntetizujícího se řetězce, působí jako terminátor reakce
14 Replikace DNA s normálními deoxynukleotidy Původní sekvence Denaturovaný templátový řetězec 3 5 Prfektní kopie
15 Co se stane pokud při reakci použijete směs dgtp a ddgtp?
16 Původní sekvence Templátový řetězec 3 5 Perfektní kopie Směs řetězců ukončených GUANINEM při použití směsi dgtp a ddgtp
17 Enzymová metoda sekvenování DNA (Sanger) DNASequencing.mov
18 seq4.mov
19 Automatické sekvenování DNA Je variantou enzymatického sekvenování DNA. Syntéza DNA probíhá v jedné reakci Ke značení produktů se používají čtyřmi různými fluorescenčními značkami označené primery dideoxyribonukleotidy
20 Asymetrická PCR pro sekvenování

21 Strategie barevných primerů

22 Strategie barevných terminátorů
 napojeného na počítač, _ který vyhodnocuje sekvenci.")
23 Princip detekce produktů Detekce produktů probíhá během elektroforézy pomocí laserového detektoru (laserem indukovaná fluorescence, LIF) napojeného na počítač, _ který vyhodnocuje sekvenci. GEL DETEKTOR + LASER Fragmenty
 - ESTEROVÉ TAMRA (černá) JOE (zelená) A ROX 5-FAM")
24 BARVY: - AMIDITOVÉ filtr žebříček HEX (černá) 6-FAM (modrá) C TAMRA TET (zelená) - ESTEROVÉ TAMRA (černá) JOE (zelená) A ROX 5-FAM (modrá)

25 Příklad fluorescenčního znační primerů



26 Soustava kapilár Genetický analyzátor ABI 3100 Vyhřívaná deska Příprava gelu Laserový detektor Rezervoár pufru Automatické nanášení vzorku


27 Izolace DNA Příprava knihovny pro Sangerovo sekvenování Fragmentace bp + Vektor Úprava konců a klonování Sestavení překrývajících se klonů
28 Příprava knihovny pro náhodné sekvenování genomů Při náhodném sekvenování genomu jsou nejdříve připraveny mechanickým stříháním genomové DNA fragmenty s optimální velikostí ( bp) a po úpravě jejich konců jsou nahodile naklonovány do vhodného vektoru za vzniku velkého počtu klonů. Ke štěpení DNA se využívá sonikace nebo pankreatická DNáza I za přítomnosti Mn 2+.
29 Předpokládá se, že celková informace o sekvenci obsažená v připravených malých klonech odpovídá původní DNA a sekvence fragmentů jednotlivých klonů se vzájemně překrývají. Pomocí univerzálních sekvenačních primerů připojujících se k vektoru poblíž klonovacího místa jsou stanoveny sekvence krátkých úseků na koncích klonovaných fragmentů (minimálně 500 bází). Stanovené sekvence jsou pak použity k uspořádání klonovaných fragmentů z jednotlivých vektorů do souvislých sekvencí genomové DNA - kontigů.
30 Uspořádané sekvenování Pro doplnění sekvence mezer zbývajících po náhodném sekvenování Pro sekvenování malých fragmentů DNA Dva odlišné přístupy: procházení primerem vyžaduje znalost sekvence, ke které se připojuje primer, který umožní prodloužení řetězce DNA-polymerázou. získaná sekvence z první reakce je použita pro návrh primeru pro další reakci a tento krok se opakuje, dokud není dosaženo kompletního stanovení sekvence. Tento proces nevyžaduje další klonování a minimalizuje stanovení nadbytečných sekvencí, ale správnost stanovené sekvence by měla být ověřena sekvenováním obou řetězců. postupné zkracování fragmentu exonukleázou a tvorba sousedících delecí
31 Hydrogensiřičitanové (bisulfite) sekvenování pro analýzu metylace cytosinu -Treatment of DNA with sodium bisulfate to convert non methylated cytosines into uracil and leaving 5-methylcytosine untouched in order to determine methylation percentage Process: Step 1 melt (denature) DNA to separate strands Step 2 treat with sodium bisulfate Step 3 amplify converted DNA with PCR Step 4 Clone PCR product Step 5 Sequence

32 Minisekvenování - stanovení SNP pomocí DNA čipu 1. varianta - Prodloužení primeru vázaného na čipu Jeden primer pro každý SNP, který genotypizujeme je imobilizován na sklíčku. K čipu jsou přidány multiplex PCR produkty, 3 fluorescenčně značené ddntps a DNA-polymeráza. Proběhne prodloužení primeru o jeden ddntp a výsledek reakce je vyhodnocen. Pozice primeru na čipu definuje, který SNP analyzujeme a fluorescence nukleotidu určuje genotyp příslušného SNP.
33 2. Varianta Alelově specifické prodloužení primeru Na sklíčku jsou imobilizovány dva alelově-specifické primery s bází na 3 -konci komplementární k oběma možným variantám nukleotidů v každém SNP. Produkty multiplex PCR jsou přepsány do mnoha kopií RNA pomocí RNApolymerázy. Molekuly RNA hybridizují k čipu a slouží jako templát pro prodloužení primeru, které je katalyzované pomocí zpětné transkriptázy Během zpětné transkripce jsou do každého produktu začleněny fluorescenčně značené dntp. Pro homozygotní genotypy je signál tvořený pouze jedním ze dvou alelověspecifických primerů kdežto u heterozygotních genotypů je signál tvořený oběma primery.
.")
34 3. Varianta Prodloužení primerů nesoucích specifickou sekvenci na 5 -konci Cyklické prodloužení primeru o jeden dideoxynukleotid je prováděno s denaturovanou DNA v roztoku za přítomnosti fluorescenčně značených ddntps, DNA-polymerázy primerů nesoucích na 5 -konci přídatnou sekvenci (tag). DNA-čip, který je komplementární k přídatným sekvencím primerů (tag array) je potom použit pro zachycení produktů cyklické minisekvenační reakce.
35 Čipy pro komparativní sekvenování pomocí hybridizace (SBH) Sekvenování zahrnuje: Hybridizaci fluorescenčně značeného fragmentu DNA ke kompletnímu DNA čipu obsahujícímu všechny kombinace (4 8 = kombinací) 8-merů nukleotidů Délka sekvence je v tomto případě max = 256 bp Výsledek hybridizace je odečten pomocí konfokálního mikroskopu Sekvence je odvozena dedukcí z hybridizujících pozicí Metoda je perspektivní, avšak limitujícím faktorem je miniaturizace společně s možností vizuální detekce Pro sekvencování 1 Mb je třeba vytvořit čip obsahující 1, možných 20-merů

36 Princip SBH - stanovení sekvence de novo
 Probíhá vývoj technik Více čtení Delší čtení Rychlejší sekvenování Nižší cena Nové aplikace v klinické oblasti, vývoj kitů cílených na určité části genomů (dědičná onemocnění nádorová")
37 Metody sekvenování nové generace (next generation sequencing) Liší se v provedení a chemii V základu se podobají v sekvenování tisíců až miliónů klonálně amplifikovaných molekul (masivní paralelní sekvenování) Probíhá vývoj technik Více čtení Delší čtení Rychlejší sekvenování Nižší cena Nové aplikace v klinické oblasti, vývoj kitů cílených na určité části genomů (dědičná onemocnění nádorová onemocnění, metagenomika, 16S rrna) Sekvenování třetí generace Pacific Biosciences Helicos Biosciences NABsys VisiGen Biotechnologies Oxford Nanophore Technologies
38 Vývoj ceny sekvenování na příkladu HGP Transformační krok Inovace Chemie reakcí Optika Fluidika Hardware Bioinformatika
39 Typy uspořádání NGS sekvenování Single Read Paired-end read Mate-pair read
40 Metody přípravy knihovny Fragmentace Enzymatická Fyzikální (sonikace, nebulizace, Hydroshear) Připojení adaptorů Amplifikace

41 Analýza velikostí fragmentů knihovny na Agilent bioanalyzátoru


42 Selekce fragmentů podle velikosti E-gel System
43 Pyrosekvenování Sekvenování se syntézou DNA v reálném čase nevyžadující elektroforézu ani separaci fragmentů Je založené na uvolnění pyrofosfátu (PPi) při enzamatické syntéze DNA (Nyren et al., 1987) 1. krok: Sekvenační primer hybridizuje k jednořetězcovému templátu a je inkubován s: DNA polymerázou ATP sulfurylázou Luciferázou Apyrázou substrátem, adenozin 5 -fosfosulfátem (APS) luciferinem 2. krok: První ze 4 dntp datp je přidán k reakci Pokud je na matrici komplementární báze, DNA polymeráza katalyzuje připojení nukleotidu k primeru Pokud na matrici není komplementární báze nukleotid bude degradován apyrázou Postupně budou přidány jeden po druhém všechny čtyři dntp Každé připojení nukleotidu je provázeno uvolněním pyrofosfátu (PPi) v množství ekvimolárním množství přidaného nukleotidu


44 3. krok: ATP sulfuryláza kvantitativně přeměňuje PPi na ATP za přítomnosti APS Vzniklý ATP umožní luciferázou zprostředkovanou konverzi luciferinu na oxyluciferin, který vytvoří světelný záblesk zaznamenaný detektorem fotonů a zobrazený jako pík na pyrogramu 4. krok: Apyráza degraduje nepřipojené dntp a nespotřebované ATP Až je degradace kompletní je přidán další dntp, který je v pořadí Proces se znovu opakuje a sekvence je odečítána z pyrogramu Namísto standardního datp je používán α thiosubstituovaný datp, který je přijímán DNA polymerázou, ale nikoli luciferázou Metoda je ve vývoji a je používána pro identifikaci jednonukleotidových polymorfizmů (SNPs)

45 Princip pyrosekvenování pyrogram
46 Pyrosekvenování (454-sekvenování) Výhody Vysoká přesnost Flexibilita a možnost paralelního zpracování velkého množství vzorků Snadná automatizace Nevyžaduje elektroforézu a značené primery Nevýhody Rychlost pyrosekvenování je cca 1 odečtená báze/min. finančně nákladná chemie

47 Klonální amplifikace knihovny emulzní PCR Příprava jednořetězcové DNA knihovny Umožňuje zpracovat různé typy vzorků genomové DNA produkty PCR BACs cdna Frakcionace dlouhých úseků na 300- to 800-bp fragmenty Vazba dvou adaptorů A a B specifických pro 3 - a 5 -konce na každý fragment Adaptor A obsahuje vazebné místo pro primer Adaptor B B obsahuje 5'-biotinovou značku, která slouží k imobilizaci DNA knihovny na mikrosféry (DNA Capture Beads) pokryté streptavidinem
 kopií Pyrosekvenování Analýza dat s využitím bioinformatických nástrojů")
48 Klonální amplifikace knihovny emulzní PCR Separace kuliček a výběr pouze těch, které obsahují navázaný jediný fragment ssdna Klonální amplifikace knihovny v emulzi emulzní PCR, amplifikační reagencie ve směsi voda-olej mikroreaktory obsahující jedinou kuličku s unikátním fragmentem DNA z knihovny - (10 7 ) kopií Pyrosekvenování Analýza dat s využitím bioinformatických nástrojů


49 Sekvenátor FLX firmy Roche Pyrosekvenování (One Bead = One Read) Kuličky se z emulze extrahují a jsou umístěny pomocí centrifugace do PicoTiterPlate, čipu z optických vláken o rozměrech mm Průměr jamek PicoTiterPlate dovoluje umístění pouze jedné kuličky průměru 28 μm Na jednom čipu je jamek Převrstvení směsí obsahující reagencie pro pyrosekvenování, enzymy jsou rovněž imobilizovány na malých mikrosférách Spodní část čipu je v optickém kontaktu s dalšími optickými vlákny napojenými na CCD senzor - zachycení až emitovaných fotonů na 1 nukleotid


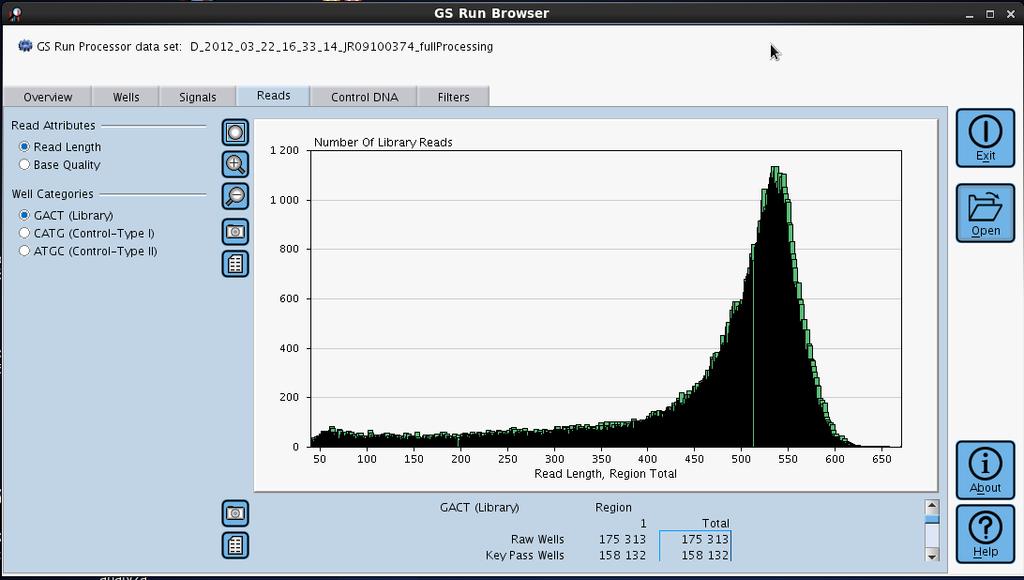
50 Sekvenátor FLX firmy Roche Kombinace vysoké intenzity signálu a pozičně specifické informace na mikrodestičce umožňuje současně analyzovat více než 1 milion sekvenačních běhů za 10 hod. Délka stanovené sekvence 400 bp Stanovení až 400 Mbp de novo Resekvenace genomů jakékoli velikosti Limitující faktory Negativní posuny čtecího rámce 3' 5' exonukleázová aktivita DNA polymerázy homopolymerní úseky Pozitivní posuny čtecího rámce Nedostatečná enzymatická aktivitá apyrázy Přesnost 99,5 % na prvních 250 bp
51 Sekvenátor FLX firmy Roche Paralelní analýza více vzorků Adaptory obsahující sekvence MID (Multiplex Identifiers) Až 12 různých unikátních sekvencí Přidány během přípravy knihovny Koamplifikovány s fragmenty DNA Rozdělení mikrodestičky do 2, 4, 8 nebo 16 menších oblastí Zabránění vzájemné kontaminace vzorků Možnost sekvenovat současně až 192 různých vzorků DNA
52
53
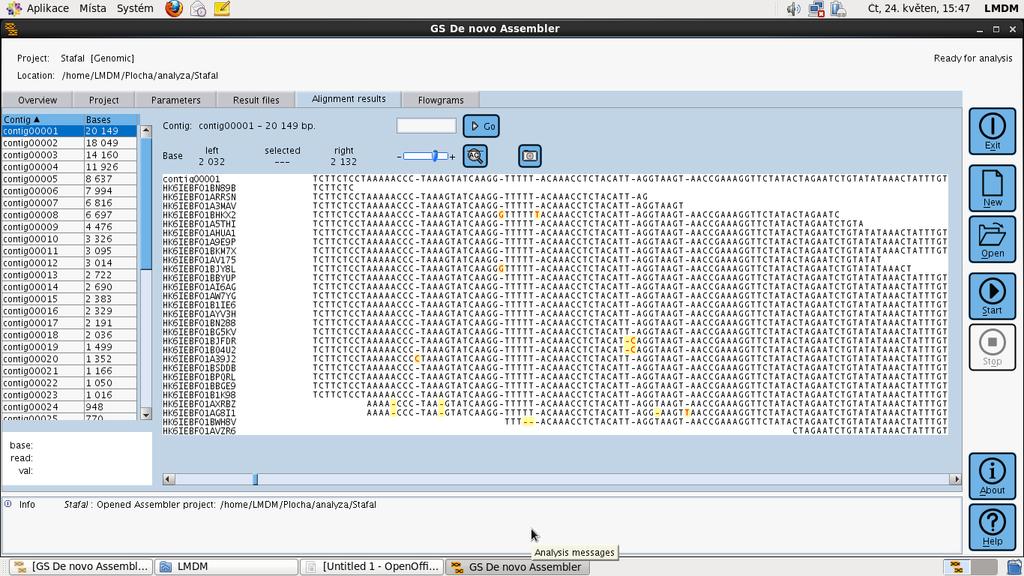
54
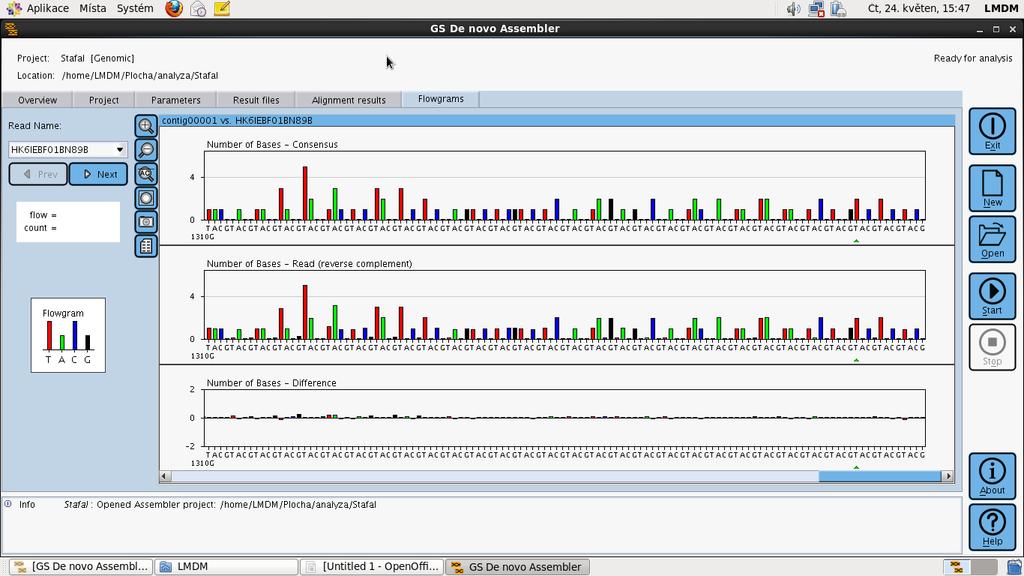
55
56 Ion Torrent polovodičové sekvenování prvním sekvenátor DNA, který nepoužívá pro detekci sekvenovaných bází DNA světelný signál, který by byl uvolňovaný při enzymatických reakcích s fluorescenčními nebo chemiluminscenčními substráty. Sekvenování Ion Torrent je založené na elektrochemické detekci vodíkových iontů (ph) které jsou uvolněny při replikaci sekvenovaného templátu na speciálním polovodičovém čipu.


57 Detekce připojené báze je založená na detekci pozitivně nabitého vodíkového iontu
58 Uvolnění vodíku v nepufrujících podmínkách
59 IonTorrent polovodičové sekvenování Na čipu se nachází milióny jednotlivých reakčních cell (reaktorů), ve kterých probíhá sekvenační analýza individuálních kopií DNA na základě kontinuálního střídání reakčních složek (jednotlivých typů nukleotidů) v mikrofluidním systému. DNA imobilizovaná na mikrosférách (emulzní PCR) Kapacita závisí na typu použitého polovodičového chipu miliónů miliónů > 1000 miliónů
60 Ion Torrent polovodičové sekvenování Technologie Ion Torrentu je otevřená pro sekvenování DNA de-novo i cílené sekvenování vybraných oblastí genomu Minimální délka sekvenačního čtení V jedno směru: 400bp Obousměrné (Paired end) Počet vzorků analyzovaných v jednom běhu až 96 vzorků pomocí označení tzv. barkodem Doba sekvenační analýzy 2-3,5 hodiny podle použitého čipu

61 Ion PGM System Sequencing Run Time 200-base reads 400-base reads 200-base reads Output* 400-base reads Ion 314 Chip v2 2.3 hrs 3.7 hrs Mb Mb Reads thousand Ion 316 Chip v2 3.0 hrs 4.9 hrs Mb 600 Mb 1 Gb 2-3 million Ion 318 Chip v2 4.4 hrs 7.3 hrs 600 Mb 1 Gb Gb million

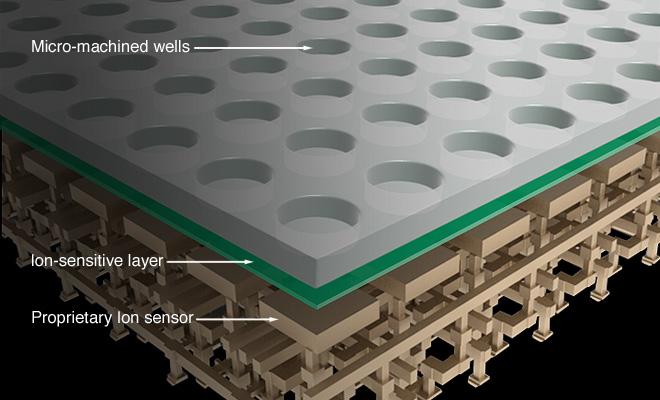
62 IonTorrent čip


63 IonTorrent - detekce dntp H + ph Q Sensing Layer Sensor Plate V Bulk Drain Source Silicon Substrate To column receiver Rothberg J.M. et al Nature doi: /nature10242 Přímá detekce iontů vodíku a konverze na elektrický signál zpracovaný čipem
64 Ionogram ACTGGATGCGGG
65 Výstup z Torrent server
66 Technologie firmy Illumina/Solexa Uvedena na trh 2006 Masivní paralelní sekvenování milionů fragmentů DNA ( polony sequencing) Sekvenacování syntézou na čipu (flow cell), které využívá strategii reverzibilních terminátorů Nevyžaduje klonování ani přípravu knihovny na mikrosférách Dosahuje 99,9 % přesnosti
 Navázání adaptorů umožňujících kovalentní vazbu na opticky transparentní povrch pro sekvenování Každý fragment je na povrchu uchycen pouze jedním")

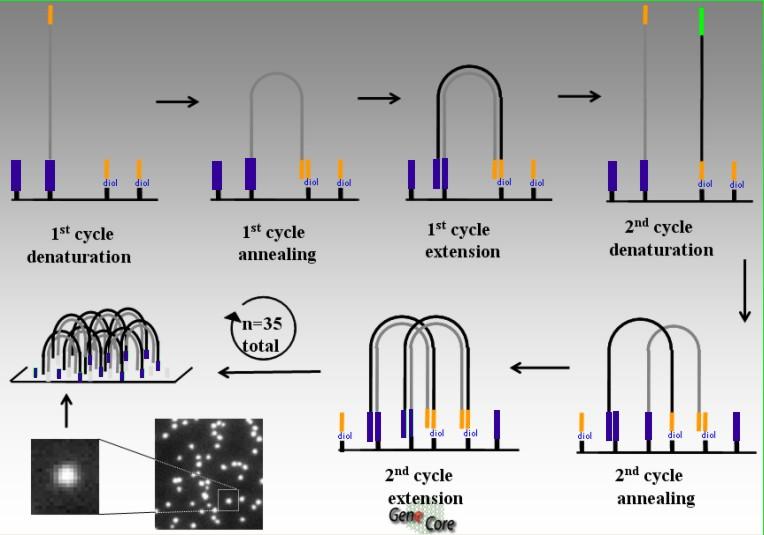
67 Illumina/Solexa Příprava knihovny Náhodná fragmentace DNA na krátké úseky bp Zatupení konců a vytvoření 1 nt A-přesahu na 3 -konci (T4 DNA polymeráza, Klenow a polynukleotid-kináza T4) Navázání adaptorů umožňujících kovalentní vazbu na opticky transparentní povrch pro sekvenování Každý fragment je na povrchu uchycen pouze jedním koncem, po přidání enzymů pro amplifikaci dochází k ohnutí fragmentu do mostu. Výsledkem amplifikace jsou dva řetězce, každý s jedním volným a jedním pevným koncem Po denaturaci jsou fragmenty narovnány auspořádány do shluků, ve kterých je dosažena značná hustota, až 1000 kopií fragmentu na μm 2 povrchu Na celém povrchu je dosaženo hustoty deseti milionů shluků na cm 2

68 Bridge amplification: initiation On the surface: complementary oligos
69

70 Illumina/Solexa Průběh sekvenační reakce Přidání primerů ve směsi s DNA polymerázou a fluorescenčně značenými dntp Nukleotidy jsou na 3'-konci modifikovány tak, že umožňují reverzibilní ukončení prodlužujícího se řetězce DNA Je tak zajištěno, že se řetězec v každém cyklu prodlouží právě o jednu bázi. Po zachycení obrazu připojené báze následuje odblokování 3'-konce

71 Illumina (Solexa) sequencing Mate-pair sequencing

72 Illumina/Solexa Délka čtených úseků: HiSeq : 100 nt or 150 nt MiSeq : 250 nt Mnohonásobné pokrytí sekvence (30 x 100 x) Možnost mnohonásobného sekvenování Povrch sklíčka rozdělen do částí Vzorky označeny pomocí dvanácti různých oligonukleotidů Současně lze analyzovat až 96 různých vzorků. Produkuje desítky až stovky Gb za 1 běh (2-8 dnů) Aplikace Resekvenování Sekvenování RNA Stanovení metylací MiSeq
73 SOLiD (Sequencing by Oligonucleotide Ligation and Detection) Nová generace vysoce účinného sekvenování, dostupná od 2008 (Applied Biosystems) Založená na kombinaci metod MPSS (Massively Parallel Signature Sequencing) [Brenner et al., 2000] Multiplex Polony Sequencing [Shendure et al., 2005] Sekvenčně specifická ligace oktamerních oligonukleotidových sond označených fluorescenčními barvivy Průměrná délka sekvenovaného úseku je 30 až 40 bp Přesnost vyšší než 99,95 % na prvních 25 bp
74 SOLiD Příprava jednoho ze dvou typů knihovny DNA Fragmentová knihovna (fragment library) Párová knihovna (mate-paired library)
75 SOLiD Amplifikace knihovny PCR v emulzi DNA knihovny Primery P1 a P2 magnetické kuličky o velikosti 1 μm vázající DNA DNA polymeráza a nukleotidy

76 SOLiD Amplifikované templáty vázané na kuličkách Denaturace templátů vázaných na kuličkách a odseparování nepoužitých kuliček

77 SOLiD Modifikace 3'-konců pro kovalentní vazbu na sklíčko pro sekvenování Nanesení kuliček Sklíčko je rozděleno na pole umožňující oddělení vzorků do jedné, čtyř nebo osmi oblastí. Klíčovou výhodou systému je možnost přípravy sklíček s velmi vysokou hustotou kuliček
78 SOLiD Sekvenování prostřednictvím ligace K adaptoru P1 vázajícímu se na kuličku hybridizuje 5 -fosforylovaný primer Je přidána sada 4 fluorescenčně značených oktamerních sond na 5'- konci Dvoubázové sondy soutěží o ligaci se sekvenačním primerem Čtyři různá barviva označují šestnáct možných kombinací na prvních dvou pozicích templátu. Specifita každé sondy v ligační reakci je podmíněna párováním první i druhé báze na 3 -konci sondy Pokud jsou první dva nukleotidy sondy komplementární s prvními dvěmi nukleotidy templátu, je sonda pomocí DNA ligázy kovalentně napojena na primer a čtecím zařízením je zachycen barevný fluorescenční signál vydaný sondou. Fosfatáza odštěpuje fosfátovou skupinu na 5'-konci prodlužovaného řetězce a tím zabraňuje vzniku fosfodiesterové vazby mezi posledním nukleotidem navázané sondy a další sondou. Dochází k terminaci prodlužujícího se řetězce. Sonda se štěpí mezi pátou a šestou pozicí, a tím dochází k odstranění a odmytí fluorescenční značky
79 Opakuje se několik cyklů (ligace, detekce, štěpení); počet cyklů určuje délku stanovené sekvence Po několika cyklech (7) je prodloužený produkt odstraněn a templát je resetován s primerem komplementárním k pozici n-1, celkem je použito 5 primerů (n, n-1, n-2, n-3 a n-4) Každá báze je integrována ve dvou nezávislých ligacích s použitím dvou primerů

80 SOLiD čtení sekvence
 Dvě odlišné dvojice bází, které mají stejnou počáteční bázi, se barví odlišnou barvou: barva (AC) barva (AG) Dvojice bází,")
81 SOLiD Pravidla a dešifrování barevného kódu Systém pracuje se 4 základními bázemi A, C, G, T. Základní barvy jsou označeny 0, 1, 2 a 3 (modrá, zelená, žlutá a červená) Dvě odlišné dvojice bází, které mají stejnou počáteční bázi, se barví odlišnou barvou: barva (AC) barva (AG) Dvojice bází, které jsou reverzní, se barví stejnou barvou: barva (AC) = barva (CA) Dvojice stejných bází se barví stejnou barvou: barva (AA) = barva (CC) Dvojice bází, které jsou odlišné, ale mají stejnou druhou bázi, se barví odlišně: barva (AC) barva (GC)
82 SOLiD paralelní analýza více vzorků SOLiD využívá šestnáct různých kódů (barcoding) pro odlišení vzorků. Vázány na 3'-konec sekvence pomocí modifikovaného adaptoru P2 Podobná teplota Tm Nízké procento chyb Unikátní v barevném kódu Možnost současně analyzovat až 256 vzorků (při využití dvou sekvenačních destiček rozdělených na osm polí). Metoda SOLiD poskytuje 9000 Mbp během 5 dnů
83 Sekvenování třetí generace Analýza jedné molekuly Potřeba malého množství vzorku Bez rizika kontaminace Přesné čtení homopolymerních úseků Analýza jakékoli NK (DNA/RNA) Analýza poškozených NK (archaické, muzejní, forenzní) Analýza DNA z nekultivovatelných organismů Absolutní kvantifikace
84 PacBio Kružnicové kontinuální sekvenování SMRT templát obsahuje dvouřetězcovou oblast (inzert) na obou koncích uzavřenou jednořetězcovými vlásenkovými smyčkami Vlásenkové smyčky představují jednořetězcovou oblast, na kterou se může vázat sekvenační primer. Polymeráza prodlužuje primer z jedné vlásenkové struktury využívá jedno vlákno DNA jako templát a druhé vytěsňuje Když se polymeráza dostane zpět k 5 konci primeru, začne vytlačovat již nasyntetizovaný řetězec a pokračuje v syntéze DNA Výsledná sekvence je odvozená z obou vláken

85 Příprava knihovny pro PacBio

86 Sekvenování třetí generace PacBio RS
87 PacBio


88 Pacific Biosciences (Roche) Sequel system PacBio RS II

89 PacBio detekce modifikovaných bází
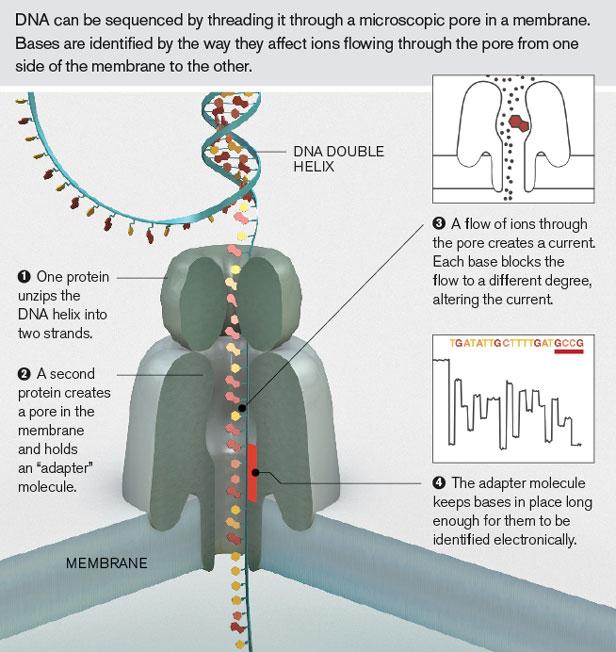
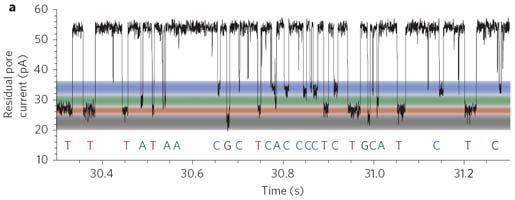
90 NanoPore (1995) Elektricky odolná polymerní membrána (sysntetická lipidová dvouvrstva) s transmembránovým porózním proteinem Modifikovaný α hemolysin (αhl) nebo Mycobacterium smegmatis porin A (MspA) Nanopore je ponořený ve vodivém roztoku Využívá měření elektrického potenciálu přes membránu Průchod bází na sekvenovaném řetězci přerušuje proud, který je specifický podle procházející báze
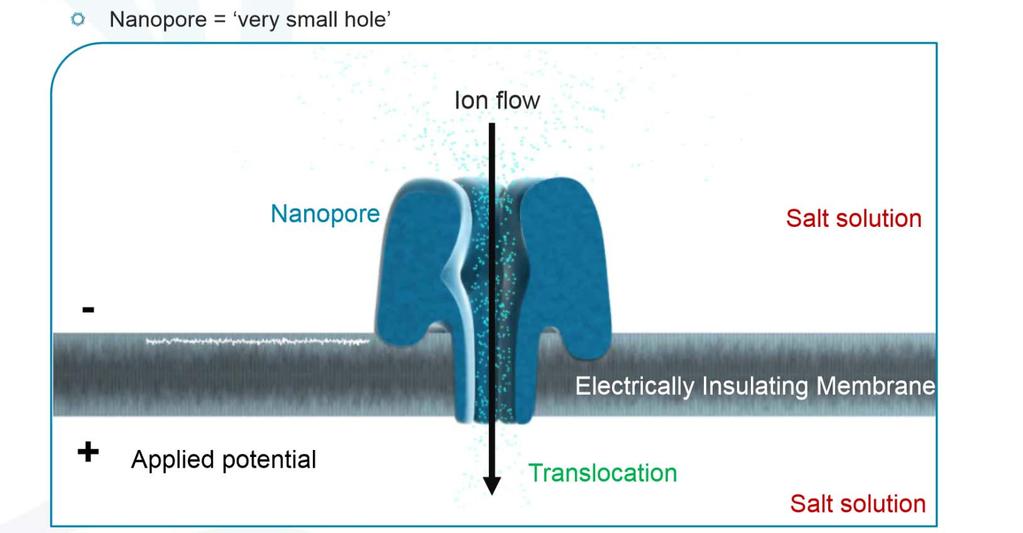
91 NanoPore


92 the MinION device the MinION device
93 Příprava knihovny MinION
94

95 Sekvenování třetí generace - Helicos
96 Srovnání metod pro sekvenování Klasické NGS Třetí generace Metoda Maxam-Gilbert (chemická) Sanger (enzymová) 454 (pyrosekvenování) Polovodičové (IonTorrent) Illumina Solid PacBio Nanopore Helicos Příprava knihovny Radioaktivně značené ssdna Inzerty v plazmidových vektorech / produkty PCR Emulzní PCR Emulzní PCR Polony PCR Emulzní PCR + imobilizace Ligace adaptorů se smyčkou Ligace adaptorů + molekulární motor Ligace adaptorů
97 Srovnání metod pro sekvenování Klasické NGS Třetí generace Metoda Maxam-Gilbert (chemická) Sanger (enzymová) 454 (pyrosekvenování) Polovodičové (IonTorrent) Illumina Solid PacBio Nanopore Helicos Princip reakce Degradace chemickými činidly Syntéza DNA polymerázou s terminátory Syntéza DNA polymerázou, detekce PP Syntéza DNA polymerázou, detekce H + Syntéza DNA polymerázou s reverzibilními terminátory Série ligací sond + posun rámce Syntéza DNA polymerázou Měření změn el. potenciálu při průchodu inertní membránou Syntéza DNA polymerázou s reverzibilními terminátory
98 Srovnání metod pro sekvenování Klasické NGS Třetí generace Metoda Maxam-Gilbert (chemická) Sanger (enzymová) 454 (pyrosekvenování) Polovodičové (IonTorrent) Illumina Solid PacBio Nanopore Helicos Délka čtení nt nt 500 nt 400 nt 200 nt nt kb > 10 kb 200 nt
99 Srovnání metod pro sekvenování Klasické NGS Třetí generace Metoda Maxam-Gilbert (chemická) Sanger (enzymová) 454 (pyrosekvenování) Polovodičové (IonTorrent) Illumina Solid PacBio Nanopore Helicos Uspořádání Single read Pair-end read Single read, Pair-end read Single read, Pair-end read Single read, Mate-pair read Mate-pair read Single read Single read Single read
Sekvenování DNA. stanovení pořadí nukleotidů v molekule DNA (primární struktury)
") Sekvenování DNA stanovení pořadí nukleotidů v molekule DNA (primární struktury) Sekvencování / Sekvenování?? Sequencing / - die Sequenzierung / - Klasické techniky sekvenování 2 metody: Chemická (Maxamova-Gilbertova)
Sekvenování DNA stanovení pořadí nukleotidů v molekule DNA (primární struktury) Sekvencování / Sekvenování?? Sequencing / - die Sequenzierung / - Klasické techniky sekvenování 2 metody: Chemická (Maxamova-Gilbertova)
Sekvenování příští generace (Next Generation Sequencing, NGS)
") Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
PŘEHLED SEKVENAČNÍCH METOD
 PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
Polymorfizmy detekované. polymorfizmů (Single Nucleotide
 Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Polymorfizmy detekované speciálními metodami s vysokou rozlišovací schopností Stanovení jednonukleotidových polymorfizmů (Single Nucleotide Polymorphisms - SNPs) Příklad jednonukleotidových polymorfizmů
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Amplifikační metody umožňují detekovat. k dispozici minimálně kopií DNA,
 Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
Diagnostické amplifikační metody nevyužívající PCR Amplifikační metody umožňují detekovat jedinou kopii cílové DNA, zatímco při hybridizačních metodách musí být k dispozici minimálně 10 4-10 5 kopií DNA,
Bi5130 Základy práce s lidskou adna
 Bi5130 Základy práce s lidskou adna Mgr. et Mgr. Kristýna Brzobohatá pizova@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu Bi5130 Základy práce s lidskou adna PCR polymerase
Bi5130 Základy práce s lidskou adna Mgr. et Mgr. Kristýna Brzobohatá pizova@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu Bi5130 Základy práce s lidskou adna PCR polymerase
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Masivně paralelní sekvenování
 Boris Tichý Sdílená laboratoř Genomika Brno, 9.10.2015 Informace je uložená v DNA Informace uložená jako sekvence bazí A, C, G, T V každé lidské buňce je ~ 3 miliardy bazí = ~ 3 metry = ~ 6.6 pikogramů
Boris Tichý Sdílená laboratoř Genomika Brno, 9.10.2015 Informace je uložená v DNA Informace uložená jako sekvence bazí A, C, G, T V každé lidské buňce je ~ 3 miliardy bazí = ~ 3 metry = ~ 6.6 pikogramů
Masivně paralelní sekvenování
 Boris Tichý Sdílená laboratoř Genomika Brno, 4. 12. 2014 Informace je uložená v DNA Informace uložená jako sekvence bazí A, C, G, T V každé lidské buňce je ~ 3 miliardy bazí = ~ 3 metry = ~ 6.6 pikogramů
Boris Tichý Sdílená laboratoř Genomika Brno, 4. 12. 2014 Informace je uložená v DNA Informace uložená jako sekvence bazí A, C, G, T V každé lidské buňce je ~ 3 miliardy bazí = ~ 3 metry = ~ 6.6 pikogramů
Molekulární genetika
 Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Výzkumné centrum genomiky a proteomiky. Ústav experimentální medicíny AV ČR, v.v.i.
 Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Ivo Papoušek. Biologie 8, 2015/16
 Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Molekulární metody ve studiích kořenových systémů. Jiří Košnar, 2016
 Molekulární metody ve studiích kořenových systémů Jiří Košnar, 2016 Úvod Řešení otázek: identifikace (barcoding) a kvantifikace rostlinných druhů ve společenstvu identifikace (barcoding) a kvantifikace
Molekulární metody ve studiích kořenových systémů Jiří Košnar, 2016 Úvod Řešení otázek: identifikace (barcoding) a kvantifikace rostlinných druhů ve společenstvu identifikace (barcoding) a kvantifikace
Elektroforéza Sekvenování
 Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION
 USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva
 EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva Iva Buriánková Katedra ekologie PřF UP KLONOVÁNÍ GENŮ KLONOVÁNÍ GENŮ Klonování
EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva Iva Buriánková Katedra ekologie PřF UP KLONOVÁNÍ GENŮ KLONOVÁNÍ GENŮ Klonování
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Analýza DNA. Co zjišťujeme u DNA
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulárně biologické metody princip, popis, výstupy
 & Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
& Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
UNIVERZITA KARLOVA V PRAZE PŘÍRODOVĚDECKÁ FAKULTA. Studijní program: Biologie Studijní obor: Biologie
 UNIVERZITA KARLOVA V PRAZE PŘÍRODOVĚDECKÁ FAKULTA Studijní program: Biologie Studijní obor: Biologie Magdaléna Pavlíková Využití metod paralelního sekvenování v mikrobiologii The use of parallel sequencing
UNIVERZITA KARLOVA V PRAZE PŘÍRODOVĚDECKÁ FAKULTA Studijní program: Biologie Studijní obor: Biologie Magdaléna Pavlíková Využití metod paralelního sekvenování v mikrobiologii The use of parallel sequencing
Využití molekulárních markerů v systematice a populační biologii rostlin. 11. Next generation sequencing (NGS)
") Využití molekulárních markerů v systematice a populační biologii rostlin 11. Next generation sequencing (NGS) Next generation sequencing (NGS) první generace Sangerovo sekvenování další generace paralelní
Využití molekulárních markerů v systematice a populační biologii rostlin 11. Next generation sequencing (NGS) Next generation sequencing (NGS) první generace Sangerovo sekvenování další generace paralelní
Enzymy v molekulární biologii, RFLP. Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek
 Enzymy v molekulární biologii, RFLP Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek Enzymy v molekulární biologii umožňují nám provádět celou řadu přesně cílených manipulací Výhody enzymů:
Enzymy v molekulární biologii, RFLP Molekulární biologie v hygieně potravin 3, 2014/15, Ivo Papoušek Enzymy v molekulární biologii umožňují nám provádět celou řadu přesně cílených manipulací Výhody enzymů:
Laboratoř sekvenace DNA Servisní laboratoř biologické sekce PřF UK
 Laboratoř sekvenace DNA Založena 2003, momentálně jsme 2 zaměstnankyně, máme dvě spolupracovnice na DPP a jsme finančně soběstačné, včetně platů. Rok Počet sekvenací Počet fragmentací 2009 10 700 8 100
Laboratoř sekvenace DNA Založena 2003, momentálně jsme 2 zaměstnankyně, máme dvě spolupracovnice na DPP a jsme finančně soběstačné, včetně platů. Rok Počet sekvenací Počet fragmentací 2009 10 700 8 100
Determinanty lokalizace nukleosomů
 METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
Analýza DNA. Co zjišťujeme u DNA DNA. PCR polymerase chain reaction. Princip PCR PRINCIP METODY PCR
 o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) SNPs Odvozování a genotyping Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s problematikou hledání
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) SNPs Odvozování a genotyping Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s problematikou hledání
Metagenomika NGS (454, Illumina, IonTorrent) Petra Vídeňská, Ph.D.
 Petra Vídeňská, Ph.D.") Metagenomika NGS (454, Illumina, IonTorrent) Petra Vídeňská, Ph.D. 1 Next Generation Sequencing The total number of genes is estimated at around 30,000--much lower than previous estimates of 80,000 to
Metagenomika NGS (454, Illumina, IonTorrent) Petra Vídeňská, Ph.D. 1 Next Generation Sequencing The total number of genes is estimated at around 30,000--much lower than previous estimates of 80,000 to
SEKVENAČNÍ METODY NOVÉ GENERACE: JEJICH PRINCIPY A POTENCIÁLNÍ VYUŢITÍ V GENETICE ČLOVĚKA, ETICKÉ ASPEKTY
 BAKALÁŘSKÁ PRÁCE MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMENTÁLNÍ BIOLOGIE ODDĚLENÍ GENETIKY A MOLEKULÁRNÍ BIOLOGIE SEKVENAČNÍ METODY NOVÉ GENERACE: JEJICH PRINCIPY A POTENCIÁLNÍ VYUŢITÍ
BAKALÁŘSKÁ PRÁCE MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMENTÁLNÍ BIOLOGIE ODDĚLENÍ GENETIKY A MOLEKULÁRNÍ BIOLOGIE SEKVENAČNÍ METODY NOVÉ GENERACE: JEJICH PRINCIPY A POTENCIÁLNÍ VYUŢITÍ
Polymerázová řetězová reakce. Základní technika molekulární diagnostiky.
 Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Polymerázová řetězová reakce Základní technika molekulární diagnostiky. Kdo za to může? Kary Mullis 1983 Nobelova cena 1993 Princip PCR Polymerázová řetězová reakce (polymerase chain reaction PCR) umožňuje
Enzymy používané v molekulární biologii
 Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Vícefunkční chemické a biochemické mikrosystémy Strana 1. Mikrofluidní bioaplikace
 Vícefunkční chemické a biochemické mikrosystémy Strana 1 Mikrofluidní bioaplikace Vícefunkční chemické a biochemické mikrosystémy Strana 2 Mikrofluidní aplikace pro bioanalýzu Transport, dávkování, promíchávání
Vícefunkční chemické a biochemické mikrosystémy Strana 1 Mikrofluidní bioaplikace Vícefunkční chemické a biochemické mikrosystémy Strana 2 Mikrofluidní aplikace pro bioanalýzu Transport, dávkování, promíchávání
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 5. Metody molekulární biologie II DNA footprinting hledání interakcí DNA s proteiny Polymerázová řetězová reakce (Polymerase chain reaction PCR) Malé
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 5. Metody molekulární biologie II DNA footprinting hledání interakcí DNA s proteiny Polymerázová řetězová reakce (Polymerase chain reaction PCR) Malé
Ivo Papoušek. Biologie 6, 2017/18
 Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 6, 2017/18 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Co zjišťujeme u DNA ACGGTCGACTGCGATGAACTCCC ACGGTCGACTGCGATCAACTCCC ACGGTCGACTGCGATTTGAACTCCC
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Modifikace PCR, sekvenování. Molekulární biologie v hygieně potravin Molekulárně biologická analýza potravin Přednáška 5, 2017/18, Ivo Papoušek
 Modifikace PCR, sekvenování Molekulární biologie v hygieně potravin Molekulárně biologická analýza potravin Přednáška 5, 2017/18, Ivo Papoušek Multiplex PCR Reakční směs obsahuje ne jeden, ale několik
Modifikace PCR, sekvenování Molekulární biologie v hygieně potravin Molekulárně biologická analýza potravin Přednáška 5, 2017/18, Ivo Papoušek Multiplex PCR Reakční směs obsahuje ne jeden, ale několik
Univerzita Karlova v Praze Přírodovědecká fakulta
 Univerzita Karlova v Praze Přírodovědecká fakulta Biologie Biologie obecná Žaneta Chmelařová Sekvenování nové generace, jeho principy a aplikace pro studium lidských nádorových onemocnění Principles of
Univerzita Karlova v Praze Přírodovědecká fakulta Biologie Biologie obecná Žaneta Chmelařová Sekvenování nové generace, jeho principy a aplikace pro studium lidských nádorových onemocnění Principles of
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
NGS analýza dat. kroužek, Alena Musilová
 NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Modifikace PCR, sekvenování. Molekulární biologie v hygieně potravin 5, 2013/14, Ivo Papoušek
 Modifikace PCR, sekvenování Molekulární biologie v hygieně potravin 5, 2013/14, Ivo Papoušek Multiplex PCR Reakční směs obsahuje ne jeden, ale několik párů primerů rozpoznávajících různé cílové sekvence
Modifikace PCR, sekvenování Molekulární biologie v hygieně potravin 5, 2013/14, Ivo Papoušek Multiplex PCR Reakční směs obsahuje ne jeden, ale několik párů primerů rozpoznávajících různé cílové sekvence
Enzymy používané v molekulární biologii
 Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
Enzymy používané v molekulární biologii Rozdělení enzymů 1. Podle substrátové specifity: většina metod molekulární biologie je závislá na použití enzymů, jejichž substrátem jsou nukleové kyseliny. Tyto
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR)
") POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
POLYMERÁZOVÁ ŘETĚZOVÁ REAKCE (PCR) Polymerázová řetězová reakce (PCR, z anglického Polymerase Chain Reaction) je metoda rychlého zmnožení (amplifikace) vybraného úseku DNA. Množený (amplifikovaný) úsek
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014. Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU
 V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII. Martina Nováková, VŠCHT Praha
 MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Univerzita Karlova v Praze
 Univerzita Karlova v Praze Farmaceutická fakulta v Hradci Králové Studijní program: Zdravotnická bioanalytika Katedra biochemických věd PŘÍNOS NEXT GENERATION SEQUENCING PRO LABORATORNÍ DIAGNOSTIKU Diplomová
Univerzita Karlova v Praze Farmaceutická fakulta v Hradci Králové Studijní program: Zdravotnická bioanalytika Katedra biochemických věd PŘÍNOS NEXT GENERATION SEQUENCING PRO LABORATORNÍ DIAGNOSTIKU Diplomová
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Pokročilé biofyzikální metody v experimentální biologii
 Pokročilé biofyzikální metody v experimentální biologii Ctirad Hofr 1/1 Proč biofyzikální metody? Biofyzikální metody využívají fyzikální principy ke studiu biologických systémů Poskytují kvantitativní
Pokročilé biofyzikální metody v experimentální biologii Ctirad Hofr 1/1 Proč biofyzikální metody? Biofyzikální metody využívají fyzikální principy ke studiu biologických systémů Poskytují kvantitativní
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
velké fragmenty střední fragmenty malé fragmenty
 velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
Seminář izolačních technologií
 Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Biotechnologický kurz. III. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Moderní metody analýzy genomu
 LÉKAŘSKÁ FAKULTA MASARYKOVY UNIVERSITY Interní hematoonkologická klinika LF MU a FN Brno Centrum molekulární biologie a genové terapie Moderní metody analýzy genomu Aplikace 25.11. 2011 Boris Tichý Aplikace
LÉKAŘSKÁ FAKULTA MASARYKOVY UNIVERSITY Interní hematoonkologická klinika LF MU a FN Brno Centrum molekulární biologie a genové terapie Moderní metody analýzy genomu Aplikace 25.11. 2011 Boris Tichý Aplikace
UNIVERZITA KARLOVA 1. lékařská fakulta BAKALÁŘSKÁ PRÁCE
 UNIVERZITA KARLOVA 1. lékařská fakulta BAKALÁŘSKÁ PRÁCE Nové techniky sekvenace lidského genomu a jejich uplatnění ve studiu molekulární podstaty a diagnostiky dědičně podmíněných onemocnění Next generation
UNIVERZITA KARLOVA 1. lékařská fakulta BAKALÁŘSKÁ PRÁCE Nové techniky sekvenace lidského genomu a jejich uplatnění ve studiu molekulární podstaty a diagnostiky dědičně podmíněných onemocnění Next generation
Izolace, klonování a analýza DNA
 Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Využití rekombinantní DNA při studiu mikroorganismů
 Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Co zjišťujeme u DNA ACGGTCGACTGCGATGAACTCCC ACGGTCGACTGCGATCAACTCCC ACGGTCGACTGCGATTTGAACTCCC
 Analýza DNA Co zjišťujeme u DNA genetickou podstatu konkrétních proteinů mutace bodové, sekvenční delece/inzerce nukleotidů, chromosomové aberace (numerické, strukturální) polymorfismy konkrétní mutace,
Analýza DNA Co zjišťujeme u DNA genetickou podstatu konkrétních proteinů mutace bodové, sekvenční delece/inzerce nukleotidů, chromosomové aberace (numerické, strukturální) polymorfismy konkrétní mutace,
PRO MOLEKULÁRNÍ BIOLOGII REAGENCIE. SLEVA 10 % na nákup nad , bez DPH. Polymerázy a mastermixy GENEDIREX Nejlepší poměr cena/výkon
 REAGENCIE PRO MOLEKULÁRNÍ BIOLOGII 092018 SLEVA 10 % na nákup nad 20 000, bez DPH Polymerázy a mastermixy GENEDIREX Nejlepší poměr cena/výkon Taq DNA polymeráza a mastermix skladování a tepelná odolnost:
REAGENCIE PRO MOLEKULÁRNÍ BIOLOGII 092018 SLEVA 10 % na nákup nad 20 000, bez DPH Polymerázy a mastermixy GENEDIREX Nejlepší poměr cena/výkon Taq DNA polymeráza a mastermix skladování a tepelná odolnost:
J09 Průkaz nukleové kyseliny
 J09 Průkaz nukleové kyseliny VLLM0421c (jaro 2016) Osnova využití a metody průkazu NK PCR a její modifikace proces prokazování specifické sekvence NK 2/55 Přímé vs. nepřímé metody přímé hledáme mikroba,
J09 Průkaz nukleové kyseliny VLLM0421c (jaro 2016) Osnova využití a metody průkazu NK PCR a její modifikace proces prokazování specifické sekvence NK 2/55 Přímé vs. nepřímé metody přímé hledáme mikroba,
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Metody studia exprese mrna. jádro a genová exprese 2007
 Metody studia exprese mrna Buněčné jádro a genová exprese 2007 Aktivita genu je primárn ě vyjád ř ena jeho transkripcí-prvním krokem vedoucím k syntéze kódovaného proteinu. Cíle metod Ur č ení mno ž ství
Metody studia exprese mrna Buněčné jádro a genová exprese 2007 Aktivita genu je primárn ě vyjád ř ena jeho transkripcí-prvním krokem vedoucím k syntéze kódovaného proteinu. Cíle metod Ur č ení mno ž ství
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA BAKALÁŘSKÁ PRÁCE
 MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA BAKALÁŘSKÁ PRÁCE BRNO 2012 KRISTÝNA KLEMENTOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Sekvenování
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA BAKALÁŘSKÁ PRÁCE BRNO 2012 KRISTÝNA KLEMENTOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Sekvenování
studium množství určitého transkriptu v daném vzorku a v množství dané molekuly mrna v dané buňce a v daném
 Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Hybridizace. doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz
 Hybridizace doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Způsoby provedení hybridizace 2) Hybridizace v roztoku 3) Příprava značených sond 4) Hybridizace
Hybridizace doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Způsoby provedení hybridizace 2) Hybridizace v roztoku 3) Příprava značených sond 4) Hybridizace
2 Inkompatibilita v systému Rhesus. Upraveno z A.D.A.M.'s health encyclopedia
 2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek
 Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek") Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
Polymerázová řetězová reakce (PCR) Molekulární biologie v hygieně potravin 4, 2013/14, Ivo Papoušek Polymerázová řetězová reakce (PCR) Zavedení PCR v roce 1983 (Kary B. Mullis) Nobelova cena 1993 Metodika
Molekulární genetika zvířat 2. Antonín Stratil. Česká zemědělská univerzita v Praze
 Molekulární genetika zvířat 2 Antonín Stratil Česká zemědělská univerzita v Praze 1. Metody studia DNA a RNA - izolace DNA - izolace RNA - zjišťování koncentrace - uchovávání DNA a RNA - elektroforéza
Molekulární genetika zvířat 2 Antonín Stratil Česká zemědělská univerzita v Praze 1. Metody studia DNA a RNA - izolace DNA - izolace RNA - zjišťování koncentrace - uchovávání DNA a RNA - elektroforéza
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
TECHNIKY PCR. PCR - polymerase chain reaction -polymerázová řetězová reakce
 TECHNIKY PCR PCR - polymerase chain reaction -polymerázová řetězová reakce Přehled Molekulárně-biologický úvod, DNA struktura, replikace, DNA polymerasa Princip procesu PCR Optimalizace PCR Typy PCR Aplikace
TECHNIKY PCR PCR - polymerase chain reaction -polymerázová řetězová reakce Přehled Molekulárně-biologický úvod, DNA struktura, replikace, DNA polymerasa Princip procesu PCR Optimalizace PCR Typy PCR Aplikace
Současné metody sekvenování nukleových kyselin Bakalářská práce
 Mendelova univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Současné metody sekvenování nukleových kyselin Bakalářská práce Vedoucí práce: prof. RNDr. Aleš Knoll, Ph.D.
Mendelova univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Současné metody sekvenování nukleových kyselin Bakalářská práce Vedoucí práce: prof. RNDr. Aleš Knoll, Ph.D.
Uživatelská příručka
 PGM Barcoding Set 1-8 Navrženo pro PGM ION-TORRENT KÓD PRODUKTU: 2001 (1-8) BALEN9: 32 testů Uživatelská příručka Rev02.2015 Str. 1 Rejstřík 1. POUŽITÍ VÝROBKU 3 2. OBSAH KITU 4 3. SKLADOVÁNÍ 4 4. STABILITA
PGM Barcoding Set 1-8 Navrženo pro PGM ION-TORRENT KÓD PRODUKTU: 2001 (1-8) BALEN9: 32 testů Uživatelská příručka Rev02.2015 Str. 1 Rejstřík 1. POUŽITÍ VÝROBKU 3 2. OBSAH KITU 4 3. SKLADOVÁNÍ 4 4. STABILITA
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
5. Sekvenování, přečtení genetické informace, éra genomiky.
 5. Sekvenování, přečtení genetické informace, éra genomiky. Minulá přednáška nastínila zrod molekulární biologie a představila některé možnosti, jak pracovat s DNA - jak ji analyzovat na základě velikosti
5. Sekvenování, přečtení genetické informace, éra genomiky. Minulá přednáška nastínila zrod molekulární biologie a představila některé možnosti, jak pracovat s DNA - jak ji analyzovat na základě velikosti