Brevipalpus phoenicis je polyfágní roztoč žijící v oblasti tropů a subtropů, kde ničí citrusy, papáju, čaj, kávu atd. za miliony dolarů ročně.
|
|
|
- Renáta Marková
- před 8 lety
- Počet zobrazení:
Transkript
1 1

2 2

, nejbližší příbuzné druhy se množí také partenogenezí nebo")
3 Brevipalpus phoenicis je polyfágní roztoč žijící v oblasti tropů a subtropů, kde ničí citrusy, papáju, čaj, kávu atd. za miliony dolarů ročně. Jeho populace je složená výhradně ze samic množících se výhradně partenogeneticky (vzácně se vyskytnuvší samci nemají žádnou funkci), nejbližší příbuzné druhy se množí také partenogenezí nebo haplodiploidií. B. phoenicis má dva chromosomy a pomocí cytogenetických technik se podařilo prokázat, že tyto chromosomy nejsou homology, neboli že tento druh je haploidní. Je to tedy jediný známý případ haploidních samic u živočichů. Haploidie je s největší pravděpodobností způsobena vnitrobuněčnou bakterií, která manipuluje pohlavím hostitele směrem k samicím, přes které je přenášena do další generace. Když byli roztoči přeléčeni antibiotiky, začali se z neoplozených vajíček líhnout samečci, původním mechanismem determinace pohlaví je tedy haplodiplodie a za nepřítomnosti bakterie by určovala pohlaví B. phoenicis dodnes. Weeks et al

4 4

5 Polyploidizace je jev poměrně běžný u rostlin (velká část druhů rostlin vznikla polyploidizací), u živočichů vzácnější a přítomná jen v některých skupinách. K opakovaným polyploidizacím docházelo a dochází u ryb a obojživelníků, u ryb se také polyploidizace navozuje uměle z šlechtitelských důvodů. U člověka sice vznikají tri- a tetraploidní embrya, ale tento stav je letální. Polyploidizace funguje velmi často jako reprodukční bariéra mezi polyploidem a rodičovskou populací, takže díky ní vznikají nové druhy. 5

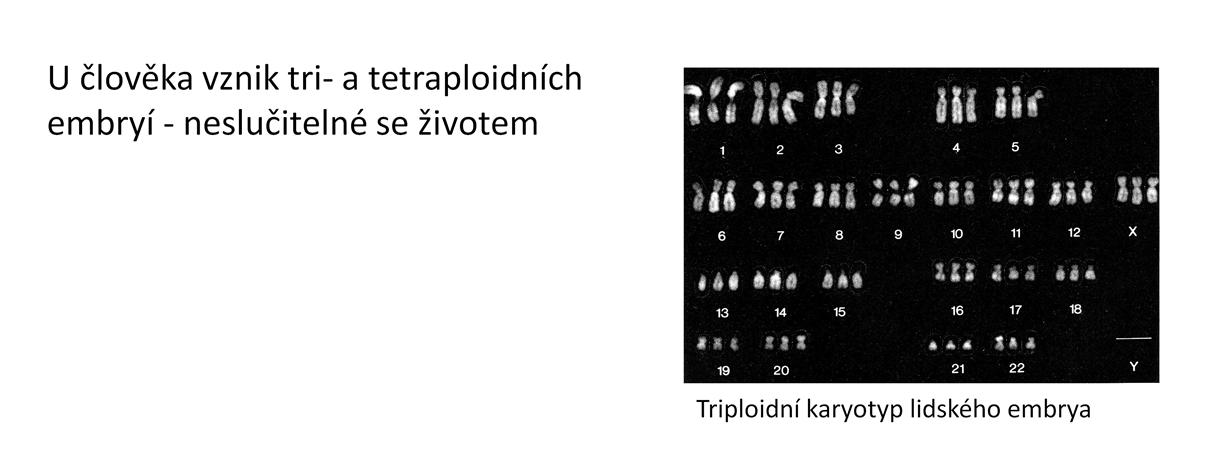
6

.")
7 Polyploidie můžeme rozlišit podle toho, zda buňka obsahuje sudý (ortoploidie) nebo lichý (anortoploidie) počet sad chromosomů. Lichý počet chromosomálních sad způsobuje problémy s rozchodem chromosomů v meióze, což vede ke sterilitě (téměř nevznikají funkční gamety). U ortoploidů je tento problém menší (nicméně také často mají problémy s meiózou), ale pokud by došlo ke křížení s diploidním jedincem, vznikne sterilní anortoploidní potomek. Je tedy žádoucí, aby docházelo k páření mezi jedinci stejné ploidie. Pokud je polyploid schopen samooplození, což je případ řady rostlin, může bez problémů produkovat potomstvo vznikl nový druh. U živočichů, kteří nemají možnost samooplození, je polyploidizace životaschopná jen pokud se polyploid dokáže množit partenogeneticky. Příkladem takového druhu je triplodiní partenogenetická ještěrka Aspidoscelis neomexicana, která je hybridem A. inortana a A. tigris. 7

8 Podle toho, zda došlo k duplikaci genomu v rámci druhu nebo při mezidruhové hybridizaci, rozlišujeme dva typy ploidií: autopolyploidie v rámci druhu, vzniklý polyploid obsahuje jen jeden typ genomu, a allopolyploidie hybridizací vznikne jedinec s různými rodičovskými genomy. Allo- i autopolyploidie způsobují ihned po vzniku masivní epigenetické změny a s tím spojenou vyšší aktivitu mobilních elementů, oba typy ploidií poskytují materiál k neofunkcionalizaci, atd. Ale allopolyploidie je zdá se výhodnější než autopolyploidie, protože dva různé genomy poskytují více alel, více proteinových interakcí, dochází k heteróznímu efektu, hybridi mají širší ekologický záběr než rodičovské druhy, vykazují vyšší rezistenci k parazitům než jeden nebo oba rodičovské druhy.

9 Většina informací, které máme o tom, co se děje bezprostředně po polyploidizaci, pochází z rostlin, kde se jednak vyskytují přirození polyploidi a jednak lze u mnoha druhů uměle polyploidy generovat. Relativně recentní duplikace genomu u obratlovců se odehrály u různých žab, ryb čeledí Salmonidae a Cyprinidae, a některých ještěrek. Přesto jsou vlivy polyploidizace u obratlovců zatím málo studované. Polyploidičasto vykazují aneuploidii, která je způsobená poruchami v meióze. Časté přestavby jsou způsobeny homeologní rekombinací spojenou s nepřesným párováním. Homeologní rekombinace je také spojena se ztrátou homeoalel u části gamet. Čím větší byly rozdíly mezi homeology, tím dramatičtější jsou následné přestavby, zahrnující inverze, translokace, tvorbu dicentrických chromosomů. Bezprostředním efektem jsou i poruchy v epigenomu, což má vliv na expresi genů a na aktivitu mobilních elementů. Geny duplikované při polyploidizaci mají vyšší naději na přežití, než geny duplikované tzv. segmentální duplikací, protože polyploidizace zachovává všechny interagující geny, zatímco duplikace menšího segmentu kopíruje jen část dráhy, jeden gen nebo jeho část. Obecně geny vzniklé segmentální duplikací vznikají častěji, ale přežívají v průměru několik milionů let, zatímco geny vzniklé polyploidizací u drápatek přežily desítky milionů let.

10 Duplikované geny může potkat řada osudů. Nejpravděpodobnější je to, že jedna z kopií je vyřazena mutací a gen je změněn na pseudogen. To může být žádoucí, pokud samostatně duplikovaný gen (vs. polyplodizace) narušuje rovnováhu genových produktů, které spolu interagují, např. v metabolické dráze. Gen může být vyřazen buď mutací v kódující oblasti (záměna nukleotidů, delece, inzerce), nebo degenerací regulačních faktorů, epigenetickou inaktivací nebo fungováním MEs. Pokud jsou zachovány obě kopie, mohou existovat obě beze změny, pokud je výhodné mít dvě kopie takového genu (např. jako pojistka proti škodlivým mutacím nebo pokud je výhodné mít více genového produktu, např. geny pro histony nebo rrna). Jedna z kopií genu může díky mutaci získat novou funkci (neofunkcionalizace). Případně může jedna nebo obě kopie ztratit některou z funkcí, které měl původní gen (subfunkcionalizace), čili dojde ke specializaci, kdy každá z kopií zastává jednu z funkcí původního genu, případně se mohou obě kopie exprimovat na různých místech nebo v jiném čase. Evans

11


12 Zvýšení odolnosti ke škůdcům a nepříznivému prostředí Navození polyploidie v některých případech vede ke zvýšení odolnosti k biotickým i abiotickým faktorům. Není to však pravidlem a někdy je efekt opačný. Výhodnější je produkce allopolyploidů z druhů, které využívají různé sekundární metabolity na obranu proti stresorům, protože sekundární metabolity jsou obvykle aditivní. Výsledný hybrid tak produkovat všechny enzymy a obrané látky, jako oba jeho rodiče. Zvětšení velikosti - Větší genom vede k větší velikosti buňky, což může být ve šlechtitelství kýženým znakem. Na některé hybridy to ale má špatný vliv, např. tetraploidi jablek jsou sice větší, ale vodnatí a špatného tvaru. Ke komerčním účelům se proto pěstují triploidi. Ploidie je žádaná i u dekorativních květin, které jsou mohutnější a déle vydrží. Zahradní jahoda je oktoploid. 12


13 Výroba sterilních kultivarů Při uvádění nových druhů do krajiny je žádoucí předejít tomu, aby se nekontrolovaně množily. Nejefektivnější metodou je navození polyploidie, která je často provázena problémy s párováním a rozchodem, takže jedinec netvoří funkční gamety. Triploidní jedinci, kteří netvoří semena, jsou žádaní v produkci ovoce. Některé druhy ale mají tzv. interploidní blok, čili jedinci různých ploidií se nekříží. To lze vyřešit kultivací triploidního endospermu in vitro, ze kterého za vhodných podmínek vznikne embryo a z něj rostlinka. Tento přístup byl úspěšně využit např. u jablek, citrusů, kiwi, mučenky, akácie, rýže a papáji. 13

14 Znovunastolení fertility u hybridů vdálených druhů Pokud jsou rodičovské druhy fylogeneticky vzdálené a tedy jejich genomy diverzifikované, jejich hybridi jsou často neplodní. Důvodem je problém s párováním homeologů a následně jejich špatný rozchod v meióze, což vede ke vzniku aberantních gamet. Zdvojením sad chromosomů se každý chromosom má s kým párovat a může se správně rozejít, takže vzniklé gamety mají správný počet chromosomů a jsou funkční. Tímto způsobem vznikla třeba Chitalpa tashkentensis (kříženec Chilopsis linearis a Catalpa bignonioides). 14

15 Případem znovunastolení fertility polyploidizací je produkt mezirodového křížení zelí a ředkvičky tzv. Raphanobrassica, Pokus prováděl ve 30. letech 20. století sovětský genetik G.D. Karpečenko a jeho cílem bylo získat rostlinu, která bude mít kořeny ředkvičky a listy zelí. Výsledek byl přesně opačný a vzniklá rostlin navíc netvořila téměř žádná semena. Z těch, která přece vznikla, vyrostly allotetraploidi, čili semena vznikla díky náhodné tetraploidizaci. 15

16 16
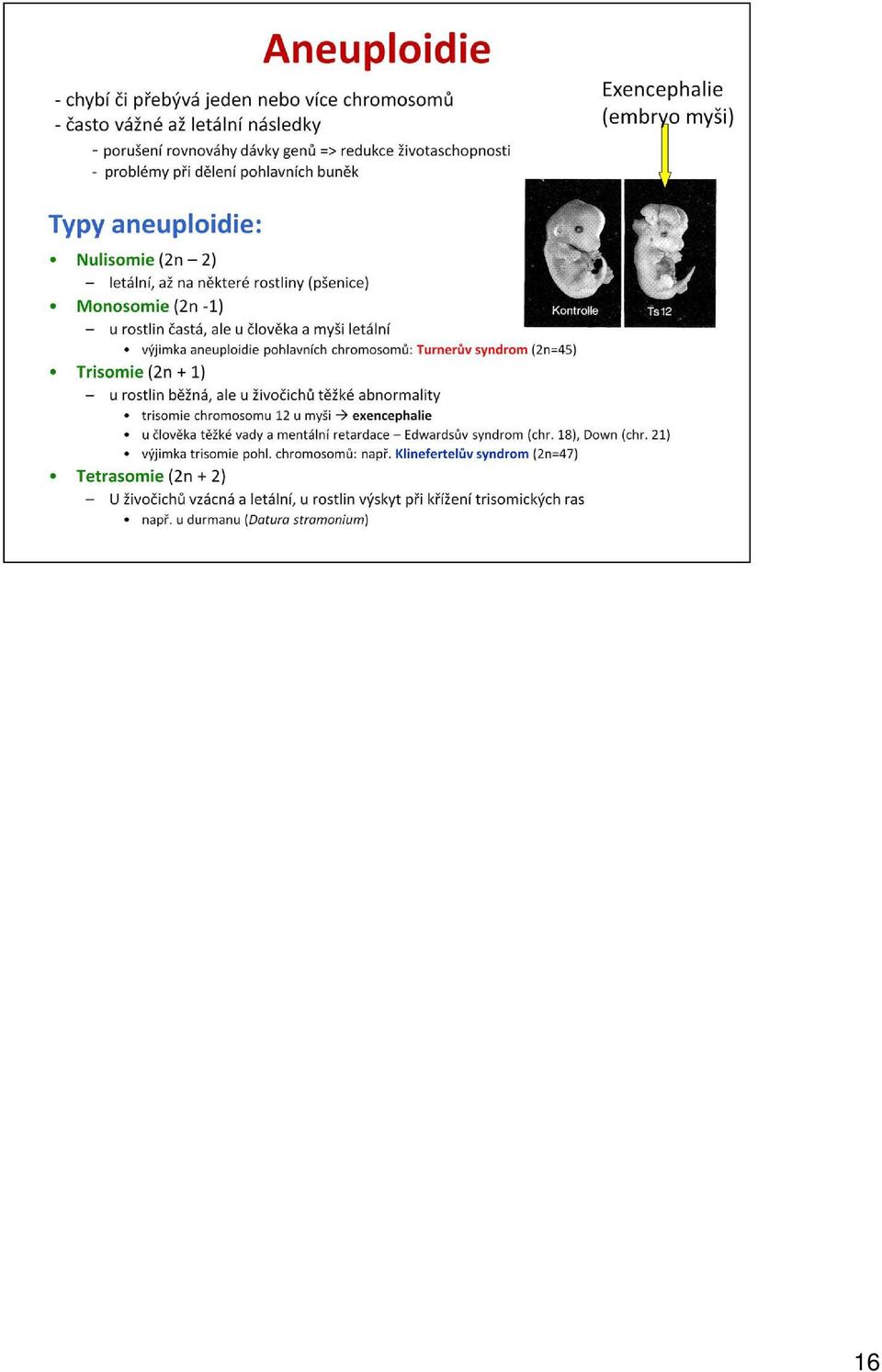
17 Aneuploidie jsou u člověka nejčastější příčinou raných potratů. Z tabulky shrnující výskyt trisomií autosomů u člověka vyplývá, že některé trisomie jsou natolik závažné, že nejsou vůbec zachytitelné ani u potracených plodů (1 a 19) a drtivá většina dalších vede k potratu. Jen trisomie 13, 18 a 21 jsou slučitelné se životem, i když trisomie 13 (Patauův syndrom) a 18 (Edwardsův syndrom) způsobují tak závažné poškození, že děti obvykle umírají krátce po porodu. Jediná trisomie, jejíž nositelé se mohou dožít dospělosti, je trisomie 21 (Downův syndrom). Sumner
 a 18 (Edwardsův syndrom) způsobují tak závažné")
18 18
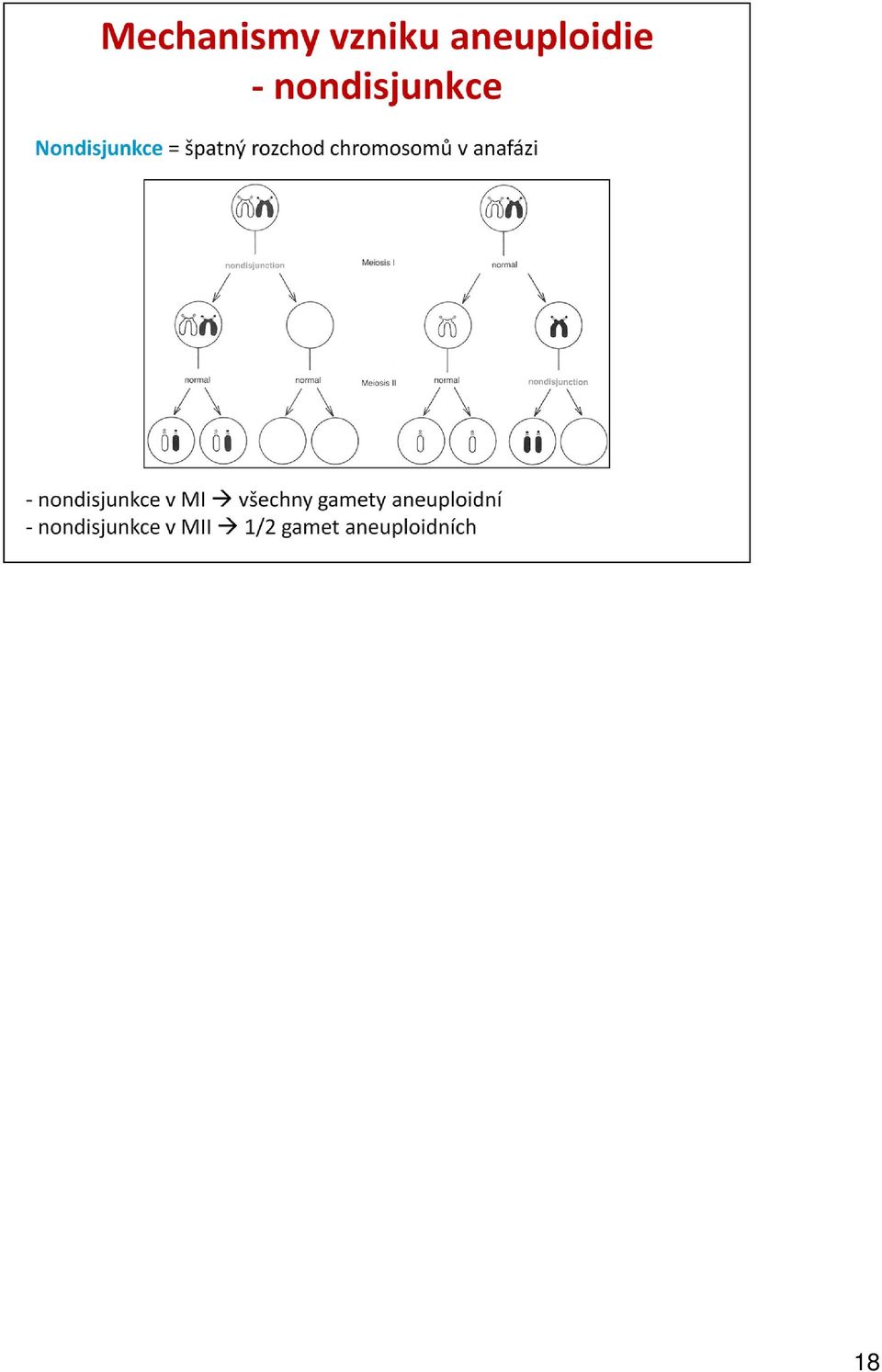
19 Robertsonovská translokace je významným jevem v evoluci karyotypu druhů. Jedná se o fúzi akrocentrických chromosomů, při které dojde k ztrátě krátkých ramének a dlouhá raménka obou akrocentriků jsou spojena v jeden chromosom s jednou centromerou. Jedinec, který je heterozygotní pro Robertsonovkou translokaci, produkuje tři druhy gamet: normální (nesou správný počet nesfúzovaných chromosomů), balancované (nese zfúzovaný chromosom) a nebalancované (část genetického materiálu jim chybí nebo přebývá). 19

20 20


21 Dvouřetězcové zlomy DNA u monocentrických chromosomů generují acentrické fragmenty (= fragmenty bez centromery), které se během metafáze nemohou zachytit na dělící vřeténko a jsou během několika generací ztraceny. Konce chromosomů bez telomer jsou lepivé a mají tendenci se spojovat, takže vnikají dicentrické chromosomy, které se v anafázi nemohou rozejít a vytvářejí tzv. anafázní můstky, což způsobí blok buněčného dělení. U holokinetických chromosomů, které mají kinetochor po celé délce/většině chromosomu, ke ztrátě fragmentů nedochází, protože se mohou navázat na dělící vřeténko. 21
22 22
23 Segmentální duplikace je zdvojení části chromosomu. Nejčastějším mechanismem je ektopická rekombinace, čili homologní rekombinace mezi úseky, které se nacházejí na různých místech chromosomu/ů (hlavně mobilními elementy). Vyskytují se ve všech genomech, i když u různých druhů jsou různě významné. Jejich prostřednictvím genom získává nový a nadbytečný genetický materiál, které může využít pro nové účely. Segmentální duplikace byly např. klíčové pro evoluci primátů. Geny v segmentálních duplikacích primátů se často účastní imunitní reakce, olfaktorického rozpoznávání, odbourávání xenobiotik a reprodukce. Naopak málo zastoupené jsou geny účastnící se běžného buněčného metabolismu a základních funkcí, patrně proto, že změna jejich exprese by byla předmětem silné purifikující selekce. Některé geny, které odlišují člověka od šimpanze vznikly duplikací. Příkladem je expanze genu pro amylázu (AMY1), který kóduje enzym z počátku metabolické dráhy škrobu a glykogenu. Tohoto genu má člověk 3x víc než šimpanz (a stejně tak i jeho produktu), což je pravděpodobně adaptace na množství potravy bohaté na škrob v zemědělských kulturách. Duplikace genu a rezistence Dobře prozkoumaným případem adaptace na insekticid duplikací genů je komár Culex pipiens, který byl na jihu Francie dlouhodobě vystavován tlaku insekticidů. Odpovědí byla duplikace genů pro dvě nespecifické esterázy Est-2 a Est-3, které odbourávají organofosfáty, a Ace1, lokus, který kóduje enzym acetylcholinesterázu, která je hlavním cílem organo-fosfátových insekticidů. 23
24 Inverze jsou další aberací, která je významná pro evoluci genomu. U heterozygotů pro inverzi mezi invertovanými úseky nedochází k párování homologů a nemůže dojít k rekombinaci, kombinace alel tak zůstávají zachované. To je důležitý mechanismus při speciaci, kdy zůstávají při obě varianty genů, které jsou např. zodpovědné za znaky odlišující vznikající druhy a za jejich rozpoznávání. Inverze jsou důležité i pro evoluci pohlavních chromosomů (viz přednáška č. 07). Obrázek z 24
25 Filadelfský chromosom je změna v karyotypu, která vzniká reciprokou translokací mezi chromosomy 9 a 22 ((t(9;22)(q34.1;q11.2)). Je typická pro chronickou myeloidní leukemii (95% pacientů), ale je přítomná i u akutní lymfoblastické leukemie (25-30% dospělých a 2-10% dětí) a příležitostně u akutní myeloidní leukemie. Translokací vzniká prodloužený chromosom 9 a zkrácený chromosom 22 (Filadelfský chromosom), který nese fúzní onkogen BCR-Abl, vzniklý spojením genu Abl1 z chromosomu 9 a část genu BCR ("breakpoint cluster region") z chromosomu 22. Gen Abl1 kóduje tyrosin kinázu, která řídí aktivitu dalších genů řídících buněčný cyklus. Onkogen BCR-Abl je transkribován kontinuálně, což vede k neregulovanému růstu buněk, čili k rakovině. Navíc inhibuje reparaci DNA, což vede k další nestabilitě genomu. K léčbě se používají inhibitory tyrosin kinázy. Označení Ph chromosomu (t(9;22)(q34.1;q11.2) znamená translokace podproužku 1 z proužku 4 z oblasti 3 z chromosomu 9 na podproužek 2 z proužku 1 z oblasti 1 chromosomu 22. (Obrázky a info z Obrázky z: 25
26 Obrázek z 26
27 Robertsonovská traslokace může být příčinou Downova syndromu, pokud k fúzi došlo mezi chromosomem 21 a dalším akrocentrikem. Sám nositel balancované translokace je v pořádku, ale část jeho gamet je nebalancovaných a nese oba chromosomy 21. Po oplození normální gametou vzniká jedinec s trisomií 21, čili s Downovým syndromem. Speciálním a vzácným případem je fúze obou chromosomů 21, kdy balancované gamety nemohou vniknout a všichni potomci balancovaného rodiče mají Downův syndrom. David-Padilla et al
28 Mutageny jsou látky, které zvyšují frekvenci mutací. Příklady chemických mutagenů analogy bazí nahrazují báze a způsobují chyby v párování mutace alkylační látky přidávají metylovou nebo etylovou skupinu k bazím chyba v párování interkalační látky vmezeření do DNA posun čtecího rámce látky deaminující báze přeměna cytosinu na uracil Fyzikální mutageny UV thyminové dimery = kovalentně spojené sousední thyminy, pokud nejsou opravené, způsobují problémy při replikaci DNA. Hlavní příčina melanomů. ionizující záření zlomy DNA Biologické mutageny mobilní elementy řada mechanismů, hlavně inserční mutageneze (vloží se do regulační/kódující sekvence genu), dále delece, duplikace,... viry inserční mutageneze
29 Wing spot test (test skvrn na křídlech) je elegantní, i když v současné době již málo využívaná metoda, jak testovat potenciální mutageny. Principem je křížení dvou čistých linií drozofil. První linie nese recesivní mutaci mwh (multiple hair wing), která způsobuje zmnožení trichomů na křídlech. Druhá linie nese recesivní mutaci flr (flare) způsobující deformované trichomy na křídlech. Oba geny jsou lokalizované na stejném chromosomu. Kříženci obou linií jsou heterozygotní pro obě recesivní mutace a mají tedy divoký fenotyp. Pokud dojde mezi homology k somatické rekombinaci, dostanou se na chromatidy stejného chromosomu mutantní alely obou genů. Po mitóze se v jedné dceřiných buňkách mohou sejít chromatidy nesoucí mutantní alely stejného genu, čili jedna buňka bude recesivní homozygot pro mwh, druhá pro flr. Potomci těchto buněk mají stejný genotyp, takže vedle sebe vzniknou dvě populace buněk, jedna s fenotypem mwh, druhá flr. Herozygoti jsou nakrmeni testovanou chemikálií a po nějaké době vyhodnoceni. Čím silnější je mutagen, tím více dvojitých skvrn se na křídle vyskytuje. Vyhodnocuje se minimálně 40 křídel ze samic, jedno křídlo má buněk. Metoda je tedy pracná, ale velmi citlivá.
30 30
31 Výměna sesterských chromatid (Sister chromatide exchange, SCE) SCE je výměna odpovídajících si částí dsdna mezi chromatidami jednoho chromosomu. K SCE dochází během S-fáze vlivem mutagenů, které interferují s replikací DNA (ionizující záření, chemikálie vážící se na DNA). Dochází k dvouřetězcovým zlomům DNA a k výměně částí sesterských chromatid. K SCE dochází přirozeně v malé míře, velké množství výměn ale ukazuje na vliv nějakého mutagenu na genom. Test na SCE se používá na testování potenciálních mutagenů a na otestování osob, které pracují v rizikových místech. K testu se používají buňky z periferní krve, které se kultivují po dobu 72 hodin (tak aby byly zastaveny v průběhu druhé mitózy) za přítomnosti bromo-deoxy-uridinu (BrdU), který je začleněn do nového řetězce DNA. Po druhé replikaci DNA obsahuje každý chromosom jednu chromatidu s jedním řetězcem s BrdU a jedním původním řetězcem DNA bez BrdU (ta se bude Giemsou barvit sytě) a jednou chromatidou s oběma řetězci DNA a BrdU (ta se bude barvit světle). Buněčné dělení je zastaveno kolchicinem, aby bylo co nejvíce buněk v metafázi, připraví se chromosomální preparát a obarví se Giemsou. Chromosomy, u kterých došlo k výměně sesterských chromatid, budou mít části chromatid světlé a jiné tmavé. Obrázek z Info z: adinstitute.org/blog/visualizing-genome-disarray 31
32 32
Proměnlivost organismu. Mgr. Aleš RUDA
 Proměnlivost organismu Mgr. Aleš RUDA Faktory variability organismů Vnitřní = faktory vedoucí k proměnlivosti genotypu Vnější = faktory prostředí Příčiny proměnlivosti děje probíhající při meioze segregace
Proměnlivost organismu Mgr. Aleš RUDA Faktory variability organismů Vnitřní = faktory vedoucí k proměnlivosti genotypu Vnější = faktory prostředí Příčiny proměnlivosti děje probíhající při meioze segregace
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
Inovace studia molekulární a buněčné biologie. reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. Z.1.07/2.2.00/07.0354 Předmět: KBB/OPSB íl přednášky: Dokončení problematiky Molekulární podstaty genetické informace, objasnění principu replikace
Inovace studia molekulární a buněčné biologie reg. č. Z.1.07/2.2.00/07.0354 Předmět: KBB/OPSB íl přednášky: Dokončení problematiky Molekulární podstaty genetické informace, objasnění principu replikace
Mutační změny genotypu
 Mutační změny genotypu - změny genotypu: segregace, kombinace + MUTACE - náhodné změny Mutace - genové - spontánní - chromozómové - indukované (uměle vyvolané) - genomové A) Genové mutace - změna (ztráta)
Mutační změny genotypu - změny genotypu: segregace, kombinace + MUTACE - náhodné změny Mutace - genové - spontánní - chromozómové - indukované (uměle vyvolané) - genomové A) Genové mutace - změna (ztráta)
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
Cytogenetika. chromosom jádro. telomera. centomera. telomera. buňka. histony. páry bazí. dvoušroubovice DNA
 Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
Základy klinické cytogenetiky II
 Základy klinické cytogenetiky II Mgr.Hanáková TYPY CHROMOSOMOVÝCH ABERACÍ - VYŠETŘENÍ VROZENÝCH CHROMOSOMOVÝCH ABERACÍ prenatální a postnatální vyšetření - VYŠETŘENÍ ZÍSKANÝCH CHROMOSOMOVÝCH ABERACÍ (vznikajících
Základy klinické cytogenetiky II Mgr.Hanáková TYPY CHROMOSOMOVÝCH ABERACÍ - VYŠETŘENÍ VROZENÝCH CHROMOSOMOVÝCH ABERACÍ prenatální a postnatální vyšetření - VYŠETŘENÍ ZÍSKANÝCH CHROMOSOMOVÝCH ABERACÍ (vznikajících
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
Klasifikace mutací. Z hlediska lokalizace mutací v genotypu. Genové mutace. Chromozomální mutace. Genomové mutace
 Mutace Klasifikace mutací Z hlediska lokalizace mutací v genotypu Genové mutace Chromozomální mutace Genomové mutace Vznik genových mutací Tranzice pyrim. za pyrim. C na T T na C purin za purin A na G
Mutace Klasifikace mutací Z hlediska lokalizace mutací v genotypu Genové mutace Chromozomální mutace Genomové mutace Vznik genových mutací Tranzice pyrim. za pyrim. C na T T na C purin za purin A na G
Glosář - Cestina. Odchylka počtu chromozomů v jádře buňky od normy. Např. 45 nebo 47 chromozomů místo obvyklých 46. Příkladem je trizomie 21
 Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
ZÍSKANÉ CHROMOSOMOVÉ ABERACE. Vytvořilo Oddělení lékařské genetiky FN Brno
 ZÍSKANÉ CHROMOSOMOVÉ ABERACE CHROMOSOMOVÉ ABERACE (CHA) Cílem cytogenetického vyšetření je zjištění přítomnosti / nepřítomnosti chromosomových aberací (patologických chromosomových změn) TYPY ZÍSKANÝCH
ZÍSKANÉ CHROMOSOMOVÉ ABERACE CHROMOSOMOVÉ ABERACE (CHA) Cílem cytogenetického vyšetření je zjištění přítomnosti / nepřítomnosti chromosomových aberací (patologických chromosomových změn) TYPY ZÍSKANÝCH
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Sterilita: stav, kdy se páru nedaří spontánně otěhotnět i přes pravidelný nechráněný pohlavní styk po dobu jednoho roku Infertilita: stav, kdy je pár
 Sterilita: stav, kdy se páru nedaří spontánně otěhotnět i přes pravidelný nechráněný pohlavní styk po dobu jednoho roku Infertilita: stav, kdy je pár schopen spontánní koncepce, ale žena není schopna donosit
Sterilita: stav, kdy se páru nedaří spontánně otěhotnět i přes pravidelný nechráněný pohlavní styk po dobu jednoho roku Infertilita: stav, kdy je pár schopen spontánní koncepce, ale žena není schopna donosit
BIO: Genetika. Mgr. Zbyněk Houdek
 BIO: Genetika Mgr. Zbyněk Houdek Nukleové kyseliny Nukleové kyseliny = DNA, RNA - nositelky dědičné informace. Přenos dědičných znaků na potomstvo. Kódují bílkoviny. Nukleotidy - základní stavební jednotky.
BIO: Genetika Mgr. Zbyněk Houdek Nukleové kyseliny Nukleové kyseliny = DNA, RNA - nositelky dědičné informace. Přenos dědičných znaků na potomstvo. Kódují bílkoviny. Nukleotidy - základní stavební jednotky.
Mutace jako změna genetické informace a zdroj genetické variability
 Obecná genetika Mutace jako změna genetické informace a zdroj genetické variability Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt
Obecná genetika Mutace jako změna genetické informace a zdroj genetické variability Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Souhrnný test - genetika
 Souhrnný test - genetika 1. Molekuly DNA a RNA se shodují v tom, že a) jsou nositelé genetické informace, b) jsou tvořeny dvěma polynukleotidovými řetězci,, c) jsou tvořeny řetězci vzájemně spojených nukleotidů,
Souhrnný test - genetika 1. Molekuly DNA a RNA se shodují v tom, že a) jsou nositelé genetické informace, b) jsou tvořeny dvěma polynukleotidovými řetězci,, c) jsou tvořeny řetězci vzájemně spojených nukleotidů,
ZÍSKANÉ CHROMOSOMOVÉ ABERACE. Vytvořilo Oddělení lékařské genetiky FN Brno
 ZÍSKANÉ CHROMOSOMOVÉ ABERACE CHROMOSOMOVÉ ABERACE (CHA) Cílem cytogenetického vyšetření je zjištění přítomnosti / nepřítomnosti chromosomových aberací (patologických chromosomových změn) TYPY ZÍSKANÝCH
ZÍSKANÉ CHROMOSOMOVÉ ABERACE CHROMOSOMOVÉ ABERACE (CHA) Cílem cytogenetického vyšetření je zjištění přítomnosti / nepřítomnosti chromosomových aberací (patologických chromosomových změn) TYPY ZÍSKANÝCH
GENETICKÁ INFORMACE - U buněčných organismů je genetická informace uložena na CHROMOZOMECH v buněčném jádře - Chromozom je tvořen stočeným vláknem chr
 GENETIKA VĚDA, KTERÁ SE ZABÝVÁ PROJEVY DĚDIČNOSTI A PROMĚNLIVOSTI Klíčové pojmy: CHROMOZOM, ALELA, GEN, MITÓZA, MEIÓZA, GENOTYP, FENOTYP, ÚPLNÁ DOMINANCE, NEÚPLNÁ DOMINANCE, KODOMINANCE, HETEROZYGOT, HOMOZYGOT
GENETIKA VĚDA, KTERÁ SE ZABÝVÁ PROJEVY DĚDIČNOSTI A PROMĚNLIVOSTI Klíčové pojmy: CHROMOZOM, ALELA, GEN, MITÓZA, MEIÓZA, GENOTYP, FENOTYP, ÚPLNÁ DOMINANCE, NEÚPLNÁ DOMINANCE, KODOMINANCE, HETEROZYGOT, HOMOZYGOT
Genetika zvířat - MENDELU
 Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Deoxyribonukleová kyselina (DNA)
") Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Karyotyp člověka. Karyotyp soubor chromozomů v jádře buňky. Význam v genetickém poradenství ke stanovení změn ve struktuře a počtu chromozomů
 Karyotyp soubor chromozomů v jádře buňky Význam v genetickém poradenství ke stanovení změn ve struktuře a počtu chromozomů Historie: 20. léta 20. století přibližný počet chromozomů v buňce člověka 1956
Karyotyp soubor chromozomů v jádře buňky Význam v genetickém poradenství ke stanovení změn ve struktuře a počtu chromozomů Historie: 20. léta 20. století přibližný počet chromozomů v buňce člověka 1956
Populační genetika III. Radka Reifová
 Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
1.12.2009. Buněčné kultury. Kontinuální kultury
 Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace ztrácejí diferenciační
Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace ztrácejí diferenciační
21. ČLOVĚK A DĚDIČNOST, GENETICKÁ PROMĚNLIVOST
 21. ČLOVĚK A DĚDIČNOST, GENETICKÁ PROMĚNLIVOST A. Metody studia dědičnosti člověka, dědičné choroby a dispozice k chorobám, genetické poradenství B. Mutace a její typy, modifikace, příklad z genetiky člověka
21. ČLOVĚK A DĚDIČNOST, GENETICKÁ PROMĚNLIVOST A. Metody studia dědičnosti člověka, dědičné choroby a dispozice k chorobám, genetické poradenství B. Mutace a její typy, modifikace, příklad z genetiky člověka
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Buněčné kultury. Kontinuální kultury
 Buněčné kultury Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace
Buněčné kultury Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
A. chromozómy jsou rozděleny na 2 chromatidy spojené jen v místě centromery. B. vlákna dělícího vřeténka jsou připojena k chromozómům
 Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Buněčné dělení ŘÍZENÍ BUNĚČNÉHO CYKLU
 BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
Chromozomová teorie dědičnosti. KBI / GENE Mgr. Zbyněk Houdek
 Chromozomová teorie dědičnosti KBI / GENE Mgr. Zbyněk Houdek Proč octomilka a T.H. Morgan? Drosophila melanogaster ideální objekt pro genetický výzkum : Rychlý reprodukční cyklus a snadný chov v laboratorních
Chromozomová teorie dědičnosti KBI / GENE Mgr. Zbyněk Houdek Proč octomilka a T.H. Morgan? Drosophila melanogaster ideální objekt pro genetický výzkum : Rychlý reprodukční cyklus a snadný chov v laboratorních
Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
 Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Cvičení č. 8. KBI/GENE Mgr. Zbyněk Houdek
 Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Mutace, Mendelovy zákony, dědičnost autosomální a gonosomální. Mgr. Hříbková Hana Biologický ústav LF MU Kamenice 5, Brno 625 00 hribkova@med.muni.
 Mutace, Mendelovy zákony, dědičnost autosomální a gonosomální Mgr. Hříbková Hana Biologický ústav LF MU Kamenice 5, Brno 625 00 hribkova@med.muni.cz Mutace Mutace - náhodná změna v genomu organismu - spontánní
Mutace, Mendelovy zákony, dědičnost autosomální a gonosomální Mgr. Hříbková Hana Biologický ústav LF MU Kamenice 5, Brno 625 00 hribkova@med.muni.cz Mutace Mutace - náhodná změna v genomu organismu - spontánní
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Nondisjunkce v II. meiotickém dělení zygota
 2. semestr, 1. výukový týden OPAKOVÁNÍ str. 1 OPAKOVÁNÍ VYBRANÉ PŘÍKLADY letního semestru: 1. u Downova a Klinefelterova syndromu, 2. Hodnocení karyotypu s aberací, 3. Mono- a dihybridismus, 4. Vazba genů
2. semestr, 1. výukový týden OPAKOVÁNÍ str. 1 OPAKOVÁNÍ VYBRANÉ PŘÍKLADY letního semestru: 1. u Downova a Klinefelterova syndromu, 2. Hodnocení karyotypu s aberací, 3. Mono- a dihybridismus, 4. Vazba genů
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
8 cyklinů (A, B, C, D, E, F, G a H) - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů
 - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů") Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
DUM č. 4 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 4 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Morfologie a rozdělení chromozomů, homologní
projekt GML Brno Docens DUM č. 4 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Morfologie a rozdělení chromozomů, homologní
Mutace a jejich význam pro evoluci
 Mutace a jejich význam pro evoluci Ivana Doležalová Osnova přednášky: Definice mutace Mutacionalismus Mutace spontánní a idukované Mutace selekčně pozitivní, negativní a neutrální Mutage genové, chromozomové
Mutace a jejich význam pro evoluci Ivana Doležalová Osnova přednášky: Definice mutace Mutacionalismus Mutace spontánní a idukované Mutace selekčně pozitivní, negativní a neutrální Mutage genové, chromozomové
GENETIKA Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální dědičnost
 Polygenní dědičnost Multifaktoriální dědičnost") GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
II. ročník, zimní semestr 1. týden OPAKOVÁNÍ. Úvod do POPULAČNÍ GENETIKY
 II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Cytogenetické vyšetřovací metody v onkohematologii Zuzana Zemanová
 Cytogenetické vyšetřovací metody v onkohematologii Zuzana Zemanová Centrum nádorové cytogenetiky Ústav klinické biochemie a laboratorní diagnostiky VFN a 1. LF UK v Praze Klinický význam cytogenetických
Cytogenetické vyšetřovací metody v onkohematologii Zuzana Zemanová Centrum nádorové cytogenetiky Ústav klinické biochemie a laboratorní diagnostiky VFN a 1. LF UK v Praze Klinický význam cytogenetických
METODY KLASICKÉ CYTOGENETIKY. Vytvořilo Oddělení lékařské genetiky FN Brno
 METODY KLASICKÉ CYTOGENETIKY DETEKCE VROZENÝCH CHROMOSOMOVÝCH ABERACÍ Standardní postup: vyšetření metodami klasické cytogenetiky + následně metodami molekulární cytogenetiky pruhování / barvení chromosomů
METODY KLASICKÉ CYTOGENETIKY DETEKCE VROZENÝCH CHROMOSOMOVÝCH ABERACÍ Standardní postup: vyšetření metodami klasické cytogenetiky + následně metodami molekulární cytogenetiky pruhování / barvení chromosomů
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
ší šířen CYTOGENETIKA
 CYTOGENETIKA V této kapitole se budeme zabývat genetickým álem lokalizovaným v buněčném jádře v útvarech zvaných chromosomy. Morfologie chromosomů se dynamicky mění během buněčného děl; v interfázi jsou
CYTOGENETIKA V této kapitole se budeme zabývat genetickým álem lokalizovaným v buněčném jádře v útvarech zvaných chromosomy. Morfologie chromosomů se dynamicky mění během buněčného děl; v interfázi jsou
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Základní genetické pojmy
 Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Karyologie. Typy chromosomů. Chromosom. Karyotyp člověka. Chromosomy. Koncové části lineárních chromosomů - telomery
 Karyologie Typy chromosomů A telocentrický B akrocentrický C submetacentrický D metacentrický Člověk nemá typ telocentrický! Chromosom Koncové části lineárních chromosomů - telomery telomera chromosom
Karyologie Typy chromosomů A telocentrický B akrocentrický C submetacentrický D metacentrický Člověk nemá typ telocentrický! Chromosom Koncové části lineárních chromosomů - telomery telomera chromosom
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Nauka o dědičnosti a proměnlivosti
 Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
World of Plants Sources for Botanical Courses
 Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích
 Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Genetika populací. KBI / GENE Mgr. Zbyněk Houdek
 Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
GENETIKA. Dědičnost a pohlaví
 GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA
 učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince. Co bylo dřív? Slepice nebo vejce?
 REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém
 Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: Z.1.07/2.2.00/28.0032 Mutace Mutace chyby genetické informace de novo, které jsou dále předávány potomkům
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: Z.1.07/2.2.00/28.0032 Mutace Mutace chyby genetické informace de novo, které jsou dále předávány potomkům
Vliv věku rodičů při početí na zdraví dítěte
 Vliv věku rodičů při početí na zdraví dítěte Antonín Šípek Jr 1,2, Vladimír Gregor 2,3, Antonín Šípek 2,3,4 1) Ústav biologie a lékařské genetiky 1. LF UK a VFN, Praha 2) Oddělení lékařské genetiky, Thomayerova
Vliv věku rodičů při početí na zdraví dítěte Antonín Šípek Jr 1,2, Vladimír Gregor 2,3, Antonín Šípek 2,3,4 1) Ústav biologie a lékařské genetiky 1. LF UK a VFN, Praha 2) Oddělení lékařské genetiky, Thomayerova
Dědičnost a pohlaví. KBI/GENE Mgr. Zbyněk Houdek
 Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Mutageneze vznik chyby na DNA mutagen (chemická látka / záření)
") Genotoxicita - úvod Genotoxicita: toxická látka ovlivňuje genetický materiál buňky (nukleové kyseliny) Při působení vyšších koncentrací genotoxických látek dochází k přímému úhynu buněk Nižší koncentrace
Genotoxicita - úvod Genotoxicita: toxická látka ovlivňuje genetický materiál buňky (nukleové kyseliny) Při působení vyšších koncentrací genotoxických látek dochází k přímému úhynu buněk Nižší koncentrace
ZÁKLADY BAKTERIÁLNÍ GENETIKY
 Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
u párů s poruchami reprodukce
 Reprodukční genetika Možnosti genetického vyšetření u párů s poruchami reprodukce Vyšetření potenciálních dárců gamet Renata Gaillyová, LF MU 2006 Reprodukční genetika Prenatální diagnostika Preimplantační
Reprodukční genetika Možnosti genetického vyšetření u párů s poruchami reprodukce Vyšetření potenciálních dárců gamet Renata Gaillyová, LF MU 2006 Reprodukční genetika Prenatální diagnostika Preimplantační
Základy klinické cytogenetiky chromosomy
 Základy klinické cytogenetiky chromosomy Hanáková M. SHRNUTÍ PŘEDNÁŠKY chromosomy metody přípravy chromosomových preparátů, hodnocení chromosomů, metody molekulární cytogenetiky vrozené chromosomové aberace
Základy klinické cytogenetiky chromosomy Hanáková M. SHRNUTÍ PŘEDNÁŠKY chromosomy metody přípravy chromosomových preparátů, hodnocení chromosomů, metody molekulární cytogenetiky vrozené chromosomové aberace
Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost
 Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost Registrační číslo: CZ.1.07/1. 5.00/34.0084 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Sada:
Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost Registrační číslo: CZ.1.07/1. 5.00/34.0084 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Sada:
Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
 Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Základy klinické cytogenetiky I
 Základy klinické cytogenetiky I Mgr.Hanáková DEFINICE A HISTORIE klinická cytogenetika se zabývá analýzou chromosomů (jejich počtem a morfologií), jejich segregací v meióze a mitóze a vztahem mezi nálezy
Základy klinické cytogenetiky I Mgr.Hanáková DEFINICE A HISTORIE klinická cytogenetika se zabývá analýzou chromosomů (jejich počtem a morfologií), jejich segregací v meióze a mitóze a vztahem mezi nálezy
Biologie - Oktáva, 4. ročník (humanitní větev)
") - Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
- Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Slovníček genetických pojmů
 Slovníček genetických pojmů A Adenin 6-aminopurin; purinová báze, přítomná v DNA i RNA AIDS Acquired immunodeficiency syndrome syndrom získané imunodeficience, způsobený virem HIV (Human immunodeficiency
Slovníček genetických pojmů A Adenin 6-aminopurin; purinová báze, přítomná v DNA i RNA AIDS Acquired immunodeficiency syndrome syndrom získané imunodeficience, způsobený virem HIV (Human immunodeficiency
Cvičeníč. 4: Chromozómy, karyotyp a mitóza. Mgr. Zbyněk Houdek
 Cvičeníč. 4: Chromozómy, karyotyp a mitóza Mgr. Zbyněk Houdek Chromozomy Geny jsou u eukaryotických organizmů z převážnéčásti umístěny právě na chromozómech v b. jádře. Jejich velikost a tvar jsou rozmanité,
Cvičeníč. 4: Chromozómy, karyotyp a mitóza Mgr. Zbyněk Houdek Chromozomy Geny jsou u eukaryotických organizmů z převážnéčásti umístěny právě na chromozómech v b. jádře. Jejich velikost a tvar jsou rozmanité,
Hardy-Weinbergův zákon - cvičení
 Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Molekulární genetika: Základní stavební jednotkou nukleových kyselin jsou nukleotidy, které jsou tvořeny
 Otázka: Molekulární genetika, genetika buněk Předmět: Biologie Přidal(a): jeti52 Molekulární genetika: Do roku 1953 nebylo přesně známa podstata genetické informace, genů, dědičnosti,.. V roce 1953 Watson
Otázka: Molekulární genetika, genetika buněk Předmět: Biologie Přidal(a): jeti52 Molekulární genetika: Do roku 1953 nebylo přesně známa podstata genetické informace, genů, dědičnosti,.. V roce 1953 Watson
KLINICKÁ CYTOGENETIKA SEMINÁŘ
 KLINICKÁ CYTOGENETIKA SEMINÁŘ Mgr.Hanáková ODDĚLENÍ LÉKAŘSKÉ GENETIKY FN BRNO ambulance cytogenetické laboratoře laboratoře klasické cytogenetiky laboratoř prenatální cytogenetiky laboratoř postnatální
KLINICKÁ CYTOGENETIKA SEMINÁŘ Mgr.Hanáková ODDĚLENÍ LÉKAŘSKÉ GENETIKY FN BRNO ambulance cytogenetické laboratoře laboratoře klasické cytogenetiky laboratoř prenatální cytogenetiky laboratoř postnatální