Základy proteomiky 2011
|
|
|
- Ladislava Marková
- před 7 lety
- Počet zobrazení:
Transkript
1 Základy proteomiky 2011 Proč právě proteomika? Jan Hejátko
2 Základy proteomiky 2011 Schéma přednášek ze Základů proteomiky 2011 Proč právě proteomika? (Jan Hejátko) Exprese a purifikace rekombinantních proteinů (Radka Dopitová) Funkce proteinů (Lubomír Janda) Určování třírozměrných proteinových struktur metodami proteinové krystalografie (Jaromír Marek) Dvoudimenzionální elektroforéza (Hana Konečná/Jana Bursíková) Hmotnostní spektrometrie proteinů (Zbyněk Zdráhal)
3 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů analýza vztahu mezi strukturou a funkcí proteinů diferenční proteomika analýza posttranslačních modifikací
4 Základy proteomiky 2011 Proč právě proteomika? PROTEOME = PROTEins expressed by genome (konference 2-D ELFO, Siena, 1994) DNA: GENOME, HAPLOME, EPIGENOME RNA: TRANSCRIPTOME PROTEIN: ORFEOME, PROTEOME, LOCALISOME, INTERACTOME, METABOLOME, PHENOME, PHENOME: kombinace různých dat, zahrnujících fenotyp, expresní data různých (ideálně všech) genů daného organismu a proteinová data (interakce, jednotlivé vlastnosti proteinů, ) Proč vůbec studovat proteiny, když máme tolik genetických dat? (sekvence genomů, expresní profily genů, fenotypy mutantů,?) V koncovém výsledku, tedy fenotypu, se vždy projeví regulace na všech úrovních, od genu po protein a jeho modifikaci Na konci je vždy BIOLOGICKÝ PROBLÉM!!!!
 různých organizmů Získané informace lze zpracovat pomocí bioinformatiky Genome Project at")
5 Anotace genů a odhad aminokyselinových sekvencí předpokládaných proteinů Postgenomová éra a co s informacemi, které neumíme číst dnes k dispozici v databázích více než záznamů (nr databáze) různých organizmů Získané informace lze zpracovat pomocí bioinformatiky Genome Project at NCBI
6 Predikce funkce genů in silico struktura genů struktura genů promotor počátek transkripce 5 UTR počátek translace místa sestřihu stop kodon 3 UTR polyadenylační signál TATA 5 UTR ATG.ATTCATCAT ATTATCTGATATA.ATAAATAAATGCGA 3 UTR
 různých organizmů Získané informace lze zpracovat pomocí")
7 Anotace genů a odhad aminokyselinových sekvencí předpokládaných proteinů Postgenomová éra a co s informacemi, které neumíme číst dnes k dispozici v databázích více než záznamů (nr databáze) různých organizmů Získané informace lze zpracovat pomocí bioinformatiky
8 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování


9 Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Genom vs. Proteom Danaus plexippus (monarch) Gen Transkript Protein Fenotyp
10 Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování DNA: Možná analogie s textem a jeho interpretací Kdyžadoperbtabijsemdfjfwůcsaknclůsnínjxldalnxckjcnbychcxmasizdciksrdceasnanazxcnlsdlaň. Když jsem snídal srdce. Když jsem s ní-----dal bych si------srdce na dlaň. Když jsem s ní-----dal by si------srdce na dlaň. RNA: Když jsem s ní, dal bych si srdce na dlaň. Když jsem s ní, dala by by si si srdce na na dlaň. Když jsem snídal srdce. PROTEIN:
11 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět



12 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce

13 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce sestřih RNA
 přesně v místě sestřihu na 5 konci 4.")

14 Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce odchylky rozpoznávání míst sestřihu u rostlin v praxi - příklad vývojové plasticity (nejen) rostlin identifikace mutanta s bodovou mutací (tranzice G A) přesně v místě sestřihu na 5 konci 4. exonu BsmI AlwNI analýza pomocí RT PCR prokázala přítomnost fragmentu BpmI PflMI AseI PsiI SpeI BclI kratšího než by odpovídalo cdna po normálním sestřihu CT GC GAA TT ACA AA GTT GT TAT TG TCT TG ATC CT AAA TT GAA TG CTC TT GTG TT TTC TA TTT CT CCA GG AAC TG GTG AA GCT CA CTG GT GCA AA AAC AC ATG AA GCC AA GAT AA ACA TT ATT AA TGA TG TTA AT GGC AT TAT AA AGC CA GGA AG GTT AG TAG TT GTC TC CTA AC TAG TT TTG AT CAA AG TTT TA TAC CT TCA AG TGT GC T GA CG CTT AA TGT TT CAA CA ATA AC AGA AC TAG GA TTT AA CTT AC GAG AA CAC AA AAG AT AAA GA GGT CC TTG AC CAC TT CGA GT GAC CA CGT TT TTG TG TAC TT CGG TT CTA TT TGT AA TAA TT ACT AC AAT TA CCG TA ATA TT TCG GT CCT TC CAA TC ATC AA CAG AG GAT TG ATC AA AAC TA GTT TC AAA AT ATG GA AGT TC ACA CG A sekvenace tohoto fragmentu pak ukázala na alternativní sesřih s využitím nejbližšího možného místa sestřihu v exonu 4 R L V V V S. L V L I K V L Y L Q V C PDR_U1b S no splicing EXON 3 pis1 intron 1470 BsmI BpmI E L V K L T G A K T H E A K I N I I N D V N G I I K P G R PDR exon 3 ORF AlwNI PflMI AseI PsiI SpeI PstI BclI BspMI CT GC GAA TT ACA AA GTT HpaI GT TAT TG TCT TG ATC CT AAA TT GAA TG CTC TT GTG TT TTC TA TTT CT CCA GG AAC TG StuI GTG AA GCT CA CTG GT GCA AA AAC AC ATG AA GCC AA GAT AA ACA TT ATT AA TGA TG TTA AT GGC AT TAT AA AGC CA GGA AG GTT AG TAG TT GTC TC CTA ACPvuII TAG TT TTG AT CAA AG TTT TA TAC CT TCA AG TGT GC T GA CG CTT AA TGT TT CAA CA ATA AC AGA AC TAG GA TTT AA CTT AC GAG AA CAC AA AAG AT AAA GA GGT CC TTG AC CAC TT CGA GT GAC CA CGT TT TTG TG TAC TT CGG TT CTA TT TGT AA TAA TT ACT AC AAT TA CCG TA ATA TT TCG GT CCT TC CAA TC ATC AA CAG AG GAT TG ATC AA AAC TA GTT TC AAA AT ATG GA AGT TC ACA CG A existence podobných obranných mechanizmů prokázána i u jiných organizmů (např. nestabilita mutantní mrna se vznikem předčasného stopkodonu (> bp před normálním stop kodonem) TA TT CTT CT TGC TG TTG CA GGT TA ACA CT GTT GC TTG GT CCT CC TAG CT GCG GA AAA AC AAC TT TGT TA AAG GC CTT GT CTG GA AAT TT AGA AA ACA AT CTA AA GGT TC TAA TG ATG AA AGC AG TTA TA TCA TT TTC TT GTG AA GAT TT TTT TG CTG CA GCT GT GTG AA GTT TG TAC CT TTT C R L V V V S. L V L I K V L Y L Q1653 V C PDR_U1b S no splicing AT AA GAA GA ACG AC AAC GT CCA AT TGT GA CAA CG AAC CA GGA GG ATC GA CGC CT TTT TG TTG AA ACA AT TTC CG GAA CA GAC CT TTA AA TCT TT TGT TA GAT TT CCA AG ATT AC TAC TT TCG TC AAT AT AGT AA AAG AA CAC TT CTA AA AAA AC GAC GT CGA CA CAC TT CAA AC ATG GA AAA G EXON 3 pis1 intron E L V K L T G A K T H E A K I N I I N D V N G I I K P G R L F F L L L Q Lu Teukaryot, L L L G P Pviz doporučená studijní literatura, PDR exon 3 ORF Singh and Lykke-Andersen, 2003) no splicing pis1 DEL pis1 EXON 4 PstI BspMI HpaI StuI PvuII C G K T T L L K A L S G N L E N N L K TA TT CTT CT TGC TG TTG CA GGT TA ACA CT GTT GC TTG GT CCT CC TAG CT GCG GA AAA AC AAC TT TGT TA AAG GC CTT GT CTG GA AAT TT AGA AA ACA AT CTA AA GGT TC TAA TG ATG AA AGC AG TTA TA TCA TT TTC TT GTG AA GAT TT TTT TG CTG CA GCT GT GTG AA GTT TG TAC CT TTT C pis1 intron pis1 exon 4 ORF AT AA GAA GA ACG AC AAC GT CCA AT TGT GA CAA CG AAC CA GGA GG ATC GA CGC CT TTT TG TTG AA ACA AT TTC CG GAA CA GAC CT TTA AA TCT TT TGT TA GAT TT CCA AG ATT AC TAC TT TCG TC AAT AT AGT AA AAG AA CAC TT CTA AA AAA AC GAC GT CGA CA CAC TT CAA AC ATG GA AAA G GCTGTTGCAa L F F L L L Q L T L L L G P P no splicing pis1 DEL pis1 EXON pis1 intron C G K T T L L K A L S G N L E N N L K EXON 4 pis1 exon 4 ORF GCTGTTGCAa L T L L L G P P S C G K T T L L K A L S G N L E N N L K EXON 4 PDR exon 4 ORF L T L L L G P P S C G K T T L L K A L S G N L E N N L K PDR exon 4 ORF PDR_L1 PDR_L1
 a konstantní oblast (C) a lehký (L) a těžký (H) řetězec každá z V oblastí L řetězce u myší je kódována 2 subgeny (V a J) každá z V oblastí H řetězce u myší je kódována 3 subgeny")
 subgenů (místně-specifickou rekombinací) L řetězec (κ): cca 300 V sub-genů a 4 J subgeny (300 x 4 = 1200 možností) H řetězec: cca 500 V sub-genů, 4 J")
15 Mechanismy uplatňující se při vyjadřování genetické informace přeskupování subgenů při produkci protilátek Přeskupování subgenů jako specifický mechanismus při produkci protilátek protilátky variabilní oblast (V) a konstantní oblast (C) a lehký (L) a těžký (H) řetězec každá z V oblastí L řetězce u myší je kódována 2 subgeny (V a J) každá z V oblastí H řetězce u myší je kódována 3 subgeny (V, J a D) v zárodečných liniích myších B-lymfocytů dochází k tzv. kombinatorické diversifikaci (přeskupování) subgenů (místně-specifickou rekombinací) L řetězec (κ): cca 300 V sub-genů a 4 J subgeny (300 x 4 = 1200 možností) H řetězec: cca 500 V sub-genů, 4 J subgeny a 12 D subgenů (500 x 4 x 12 = možností) celkové množství kombinací u myší: cca 1200 x = 28 mil. různých V oblastí (protilátek rozpoznávající různé antigeny) antigen indukuje tzv. afinitní dozrávání mechanismem somatické hypermutace po aktivaci B-lymf. pomocnými T-lymf. dochází ke zvýšenému výskytu mutací ve V oblastech (1 mutace/v oblast/generaci, cca 1 mil. X vyšší než je obvyklé (např. u tzv. house-keeping genů) a selekci protilátek se zvýšenou afinitou k antigenu
16 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce sestřih RNA translační represe
17 Regulace genové exprese mechanismem translační represe Funkční význam sestřihu v nepřekládaných oblastech - důležitá regulační součást genů Translační represe prostřednictvím krátkých ORF v 5 UTR Identifikováno např. u kukuřice (Wang and Wessler, 1998, viz doporučená lit.) M K R A F. ATGaaaagagcttttTAG ATGatggtgaaagttaca. V případě CKI1 pokus prokázat tento způsob regulace genové exprese pomocí transgenních linií nesoucích uida pod kontrolou dvou verzí promotoru, zatím nepotvrzeno M K R A F. ATGaaaagagcttttTAG M M V K V T ATGatggtgaaagttaca. M M V K V T ATGatggtgaaagttaca.






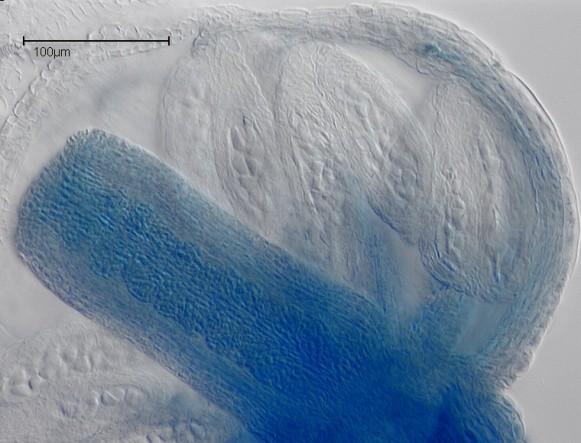
18 Expression of CKI1 in Diploid Generative Tissue Inflorescence
19 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce Sestřih RNA translační represe posttranskripční umlčování mechanizmem sirna
20 Od genu k proteinu a zpět umlčování genů mechanismem RNA interference (RNAi) Mello, 2004


21 The Nobel Prize in Physiology or Medicine 2006 "for their discovery of RNA interference - gene silencing by double-stranded RNA" Andrew Z. Fire Craig C. Mello USA Stanford University School of Medicine Stanford, CA, USA USA University of Massachusetts Medical School Worcester, MA, USA b b. 1960






 výjimkou je exprese genu AP2,")

22 Od genu k proteinu a zpět posttranskripční umlčování mechanizmem sirna Regulace vývoje květů u Arabidopsis prostřednictvím mirna ABC model vývoje květních orgánů u rostlin během vývoje květních orgánů dochází k určování identity jednotlivých květních orgánů kombinací exprese tzv. homeiotických genů homeiotické geny kódují většinou rostlinné homolgy MADS-box transkripčních faktorů mezi jednotlivými homeiotickými geny dochází k tzv. katastrálním interakcím, kdy exprese jednoho genu inhibuje expresi dalšího např. AP1 je nejprve aktivní v celém květním meristému, po indukci exprese AG pak AG inhibuje expresi AP1 ve vnitřních dvou kruzích) výjimkou je exprese genu AP2, jehož mrna je přítomná v celém květním meristému, ale exprese AP2 je regulována na úrovni translace prostředictvím mirna (gen mirna 172) wt ap2 35S::miRNA 172 in situ lokalizace mirna172 v 3. a 4. kruhu
23 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce Sestřih RNA translační represe posttranskripční umlčování mechanizmem sirna směřování proteinů


24 Intracelulární lokalizace proteinů Od genu k proteinu a zpět směřování (cílování) proteinů Pro funkci proteinů v buňkách je zásadní jejich správná lokalizace prostřednictvím tzv. signálních sekvencí v rostlinných buňkách dochází k velice dynamickým procesům, zprostředkovávaným zejména tzv. endomembránovým transportem (viz film, GFP směřované do ER) endomembránový transport je důležitým regulačním mechanismem při přenosu signálu a regulaci buněčných procesů CV, central vacuole; DV, dense vesicle; ER, endoplasmic reticulum; GA, Golgi apparatus; LV, lytic vacuole; N, nucleus; PAC, precursoraccumulating compartment; PB, protein body; PCR, partially coated reticulum; PSV, protein-storage vacuole; PVC, pre-vacuolar compartment; SV, secretory vesicle. Surpin and Raikhel, 2004.





25 Od genu k proteinu a zpět směřování (cílování) proteinů Cyklování auxinových přenašečů u Arabidopsis auxin je rostlinný hormon se silným morfogenním účinkem proteiny podílející se na transportu proteinů jsou tzv. PIN proteiny, polárně lokalizované v bunňkách kořene u Arabidopsis PIN proteiny cyklují v endomembránovém systému rostlinné buńky v přítomnosti inhibitorů endocytózy (BFA) dochází k akumulaci těchto proteinů v intracelulárních kompartmentech..čímž je zároveň negativně ovlivněn gravitropizmus u rostlin
26 Od genu k proteinu a zpět Mechanismy uplatňující se při vyjadřování genetické informace regulace transkripce sestřih RNA translační represe posttranskripční umlčování mechanizmem sirna směřování proteinů posttranslační modifikace proteinů
27 Od genu k proteinu a zpět postranslační modifikace proteinů Význam posttranslačních modifikací proteinů regulace enzymové aktivity regulace interakcí proteinu s dalšími proteiny nebo jinými biomolekulami lokalizace proteinu v buňce změna mechanických vlastností proteinu přenos signálu
28 Od genu k proteinu a zpět postranslační modifikace proteinů Typy posttranslačních modifikací proteinů přidání glykosylfosfatidylinositolové (GPI) kotvy fosforylace sulfonace glykosylace N-myristolyace N-metylace hydroxylace karboxylace prenylace.
29 Od genu k proteinu a zpět postranslační modifikace proteinů Přenos signálu a regulace genové exprese prostřednictvím fosforylace přenos cytokininového signálu u rostlin input signal P P N in p u t dom ain H is transm itter m o d u le A sp receive r m o d u le C P P output signal hybrid sensor kinase H is N C N C response regulator(s) o u tp u t dom ain

30 Signal Transduction via TCS Recent Model of the CK Signaling via TCS Pathway CYTOKININ PM AHK sensor histidine kinases AHK2 AHK3 CRE1/AHK4/WOL HPt Proteins AHP1-6 NUCLEUS Response Regulators ARR1-24 D Agostino et al., Plant Physiol, 2000 REGULATION OF TRANSCRIPTION CK primary response genes - Type-A ARRs expression INTERACTION WITH EFFECTOR PROTEINS
31 Od genu k proteinu a zpět postranslační modifikace proteinů Přenos signálu a regulace genové exprese prostřednictvím fosforylace přenos signálu prostřednictvím TGFβ (Transforming Growth Factor) u živočichů
32 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů






33 Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů Technologie rekombinatních proteinů umožňuje získat velké množství analyzovaného proteinu ve velké čistotě využívá technologie rekombinantní DNA principem je vložení přívěsku prostřednictvím přípravy rekombinantní DNA, který usnadní purifikaci (afinitní purifikace) možnost využití přívěsku i pro lokalizaci a další analýzu proteinu v in vivo systémech
34 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů analýza vztahu mezi strukturou a funkcí proteinů



35 Přístupy současné proteomiky analýza vztahu mezi strukturou a funkcí proteinů Analýza vztahu mezi funkcí a strukturou proteinu využívá technologie produkce rekombinantních proteinů a místně řízené mutageneze umožňuje analyzovat strukturu rekombinantního proteinu pomocí rentgenové krystalografie nebo NMR kopmparativní analýzou lze pak analyzovat strukturu a funkci jak u strandardního typu tak i mutantního proteinu
36 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů analýza vztahu mezi strukturou a funkcí proteinů diferenční proteomika

37 Přístupy současné proteomiky komparativní proteomika Analýza změn proteomu organismu na různé stimuly využívá technologií vícerozměrné separace proteinů (nejčastěji ELFO nebo LC) umožňuje analyzovat mnoho proteinů najednou pomocí analýzy obrazu lze odhadnout proteiny se změnou vlastností, příp. kvantity za daných sledovaných podmínek (mutace, vývojové stadium, fyziologická odpověď) Zvýšená hladina endogenních CK
38 Přístupy současné proteomiky komparativní proteomika Wt.col. O pop-ipt11(5)/lhgr pop-ipt13(2)/lhgr MS HPCd DEX
39 Přístupy současné proteomiky komparativní proteomika S pot % V ol R elative A bunda nce A ccession N o. P rotein N am e M A LD I-MS LC-MSMS M M [kd a] S core % C ov S core % C ov pi S protein hom olog R C I < 0.01 B A B A P N ID (jasm o nate iducible, m yrosinase-like) 2310 < 0.01 B A A A P N ID (R ub isc O large sub unit) Q 8LB J7 50S ribosom al protein L12-C A A G A C N ID (2-cys pero xiredoxin B A S 1, chloroplast [P rec ursor]) 4503 > 100 Q 9S U I9 A TP synthase g am m a chain, chloroplast 5204 < 0.01 E H ypothetical protein (putative epoxide hydrolase) 7009 < 0.01 T05413 cinnam yl-alcohol dehydrog e nase (E C ) F 28A < 0.01 Q 9LJE 4 G loe L protein, chapero nin, 60 kd a 7606 > 100 F86262 F13K protein (glyceraldehyde-3-p hosp ha te d h) A A M A Y N ID (C yste ine synthase, m itocho ndrial [P recursor]) 8202 > 100 A A L36245 A Y N ID (50S ribosom al protein L4, chloroplast [P recursor]) H probable anne xin > 100 Q 941D 3 A t5g19940/f28 I16_90 (H ypothetical protein) T47470 isovaleryl-c oa dehydroge nase
40 Základy proteomiky 2011 Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů analýza vztahu mezi strukturou a funkcí proteinů diferenční proteomika analýza posttranslačních modifikací


41 Přístupy současné proteomiky analýza posttrranslačních modifikací Analýza posttranslačních modifikací pomocí specifických metod lze identifikovat kotranslační a posttransalční modifikace, buď v gelu nebo po blotování na membránu (barvení spec. barvičkami) identifikace modifikací pomocí MS technik (MALDI TOF, ESI-MS, ) fosforylace přenos signálu acetylace regulace chromatinových strruktur a transkripční aktivity prostřednictvím regulace vazby histonů glykosylace velice heterogenní (aktivátor plasminogenu 3 místa pro glykosylaci na N-konci, až SYPRO možných Rubyizoforem) Pro-Q Diamond mikroheterogenita díky velkému množství různých cukerných zbytků, které se mohou vázat na jednu ak. makroheterogenita díky rozdílům v přítomnosti různých cukrů na různých ak. na různém počtu kopií daného proteinu
42 Základy proteomiky 2011 shrnutí Proč právě proteomika? Postgenomová éra a co s informacemi, které neumíme číst Genotyp vs. fenotyp, aneb co všechno se děje při vyjadřování Od genu k proteinu a zpět Přístupy současné proteomiky exprese, purifikace a analýza rekombinantních proteinů analýza vztahu mezi strukturou a funkcí proteinů diferenční proteomika analýza posttranslačních modifikací
43 Základy proteomiky 2011 zdrojová literatura Zdrojová literatura k první přednášce: Monografie a učebnice Plant Functional Genomics, ed. Erich Grotewold, 2003, Humana Press, Totowa, New Jersey Proteome Research: New Frontiers in Functioonal Genomics, ed. Wilkins, M.R., Wiliams, K.L., Appel, R.D., Hochstrasser, D.F., 1997, Springer-Verlag, Berlin, Heidelberg Dubová J., Hejátko J., Friml J. (2005) Reproduction of Plants, in Encyclopedia of Molecular Cell Biology and Molecular Medicine (ed, R. A. Meyers), pp Wiley-VCH, Weinheim, Germany Publikace v mezinárodních časopisech Wang, L. and Wessler, S.R. (1998) Inefficient reinitiation is responsible for upstream open reading frame-mediated translational repression of the maize R gene. Plant Cell, 10, (1733) Friml, J. and Palme, K. (2002) Polar auxin transport. Old questions and new concepts?. Plant Mol. Biol., 49, Mello, C.C. and Conte Jr., D. (2004) Revealing the world of RNA interference. Nature, 431, Surpin, M. and Raikhel, N. (2004) Traffic jams affect plant development and signal transduction. Nature Reviews/Molecular Cell Biology 5,
Základy proteomiky. Proč právě proteomika? Jan Hejátko
 Základy proteomiky Proč právě proteomika? Jan Hejátko Základy proteomiky zdrojová literatura Zdrojová literatura k první přednášce: Monografie a učebnice Plant Functional Genomics, ed. Erich Grotewold,
Základy proteomiky Proč právě proteomika? Jan Hejátko Základy proteomiky zdrojová literatura Zdrojová literatura k první přednášce: Monografie a učebnice Plant Functional Genomics, ed. Erich Grotewold,
Základy genomiky II, Identifikace genů
 Základy genomiky II. Identifikace genů Jan Hejátko Masarykova univerzita, Laboratoř funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky II. Zdrojová literatura ke kapitole
Základy genomiky II. Identifikace genů Jan Hejátko Masarykova univerzita, Laboratoř funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky II. Zdrojová literatura ke kapitole
Autoindex nad DNA sekvencemi
 Autoindex nd DNA sekvenemi do. Ing. Jn Holub, Ph.D. ktedr teoretiké informtiky Fkult informčníh tehnologií České vysoké učení tehniké v Prze ENBIK 2014 10. 6. 2014 ENBIK 2014, 10. 5. 2014 J. Holub: Autoindex
Autoindex nd DNA sekvenemi do. Ing. Jn Holub, Ph.D. ktedr teoretiké informtiky Fkult informčníh tehnologií České vysoké učení tehniké v Prze ENBIK 2014 10. 6. 2014 ENBIK 2014, 10. 5. 2014 J. Holub: Autoindex
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Molekulární genetika IV zimní semestr 6. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika IV zimní semestr 6. výukový týden (5.11. 9.11.2007) Nondisjunkce u Downova syndromu 2 Tři rodokmeny rodin s dětmi postiženými
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika IV zimní semestr 6. výukový týden (5.11. 9.11.2007) Nondisjunkce u Downova syndromu 2 Tři rodokmeny rodin s dětmi postiženými
Schéma průběhu transkripce
 Molekulární základy genetiky PROTEOSYNTÉZA A GENETICKÝ KÓD Proteosyntéza je složitý proces tvorby bílkovin, který zahrnuje proces přepisu genetické informace z DNA do kratšího zápisu v informační mrna
Molekulární základy genetiky PROTEOSYNTÉZA A GENETICKÝ KÓD Proteosyntéza je složitý proces tvorby bílkovin, který zahrnuje proces přepisu genetické informace z DNA do kratšího zápisu v informační mrna
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Genetický kód. Jakmile vznikne funkční mrna, informace v ní obsažená může být ihned použita pro syntézu proteinu.
 Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
ACTIVATION OF DEHYDRIN GENES OF GERMINATE PLANTS OF BARLEY TO DROUGHT AND COLD AKTIVACE DEHYDRINOVÝCH GENŮ KLÍČNÍCH ROSTLIN JEČMENE SUCHEM A CHLADEM
 ACTIVATION OF DEHYDRIN GENES OF GERMINATE PLANTS OF BARLEY TO DROUGHT AND COLD AKTIVACE DEHYDRINOVÝCH GENŮ KLÍČNÍCH ROSTLIN JEČMENE SUCHEM A CHLADEM Dokoupilová Z., Holková L., Chloupek O. Ústav pěstování
ACTIVATION OF DEHYDRIN GENES OF GERMINATE PLANTS OF BARLEY TO DROUGHT AND COLD AKTIVACE DEHYDRINOVÝCH GENŮ KLÍČNÍCH ROSTLIN JEČMENE SUCHEM A CHLADEM Dokoupilová Z., Holková L., Chloupek O. Ústav pěstování
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
OPVK CZ.1.07/2.2.00/
 OPVK CZ.1.07/2.2.00/28.0184 Základní principy vývoje nových léčiv OCH/ZPVNL Mgr. Radim Nencka, Ph.D. ZS 2012/2013 První kroky k objevu léčiva Nobelova cena za chemii 2013 Martin Karplus Michael Levitt
OPVK CZ.1.07/2.2.00/28.0184 Základní principy vývoje nových léčiv OCH/ZPVNL Mgr. Radim Nencka, Ph.D. ZS 2012/2013 První kroky k objevu léčiva Nobelova cena za chemii 2013 Martin Karplus Michael Levitt
Polymerázová řetězová reakce
 Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Polymerázová řetězová reakce doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2013 Obsah přednášky 1) Co je to PCR, princip, jednotlivé kroky 2) Technické provedení PCR 3) Fyzikální
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Populační genetika. ) a. Populační genetika. Castle-Hardy-Weinbergova zákonitost. Platí v panmiktické populaci za předpokladu omezujících podmínek
 a. Populační genetika. Castle-Hardy-Weinbergova zákonitost. Platí v panmiktické populaci za předpokladu omezujících podmínek") Poulační genetika Poulační genetika ORGANISMUS Součást výše organizované soustavy oulace POPULACE Soubor jedinců jednoho druhu Genotyově heterogenní V určitém čase má řirozeně vymezený rostor Velký očet
Poulační genetika Poulační genetika ORGANISMUS Součást výše organizované soustavy oulace POPULACE Soubor jedinců jednoho druhu Genotyově heterogenní V určitém čase má řirozeně vymezený rostor Velký očet
Využití rep-pcr v bakteriální taxonomii
 Využití rep-pcr v bakteriální taxonomii Pavel Švec Česká sbírka mikroorganismů Přírodovědecká fakulta MU rep-pcr založeny na shlukové analýze PCR produktů získaných s primery komplementárními k rozptýleným
Využití rep-pcr v bakteriální taxonomii Pavel Švec Česká sbírka mikroorganismů Přírodovědecká fakulta MU rep-pcr založeny na shlukové analýze PCR produktů získaných s primery komplementárními k rozptýleným
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION
 USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulárn. rní genetika
 Molekulárn rní genetika Centráln lní dogma molekulárn rní biologie cesta přenosu genetické informace: DNA RNA proteiny výjimkou reverzní transkripce retrovirů: RNA DNA Chemie nukleových kyselin dusíkaté
Molekulárn rní genetika Centráln lní dogma molekulárn rní biologie cesta přenosu genetické informace: DNA RNA proteiny výjimkou reverzní transkripce retrovirů: RNA DNA Chemie nukleových kyselin dusíkaté
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
ZPŮSOB DETOXIKACE SULFIDICKÉHO YPERITU ÚČINKEM HALOGENALKANDEHALOGENÁZ
 Patentová přihláška CZ 2005 352 ZPŮSOB DETOXIKACE SULFIDICKÉHO YPERITU ÚČINKEM HALOGENALKANDEHALOGENÁZ Číslo přihlášky: CZ 2005 352 A1 Datum předložení: 3. června 2005 Abstrakt: Způsob detoxikace sulfidického
Patentová přihláška CZ 2005 352 ZPŮSOB DETOXIKACE SULFIDICKÉHO YPERITU ÚČINKEM HALOGENALKANDEHALOGENÁZ Číslo přihlášky: CZ 2005 352 A1 Datum předložení: 3. června 2005 Abstrakt: Způsob detoxikace sulfidického
Lucie Kárná, Michal Křížek, Pavel Křížek
 genetika Genetický kód z pohledu matematiky Lucie Kárná, Michal Křížek, Pavel Křížek RNDr. Lucie Kárná, Ph.D. (*1969) vystudovala obor matematická analýza na Matematickofyzikální fakultě UK a v současnosti
genetika Genetický kód z pohledu matematiky Lucie Kárná, Michal Křížek, Pavel Křížek RNDr. Lucie Kárná, Ph.D. (*1969) vystudovala obor matematická analýza na Matematickofyzikální fakultě UK a v současnosti
Replikace, transkripce a translace
 Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 5. Metody molekulární biologie II DNA footprinting hledání interakcí DNA s proteiny Polymerázová řetězová reakce (Polymerase chain reaction PCR) Malé
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 5. Metody molekulární biologie II DNA footprinting hledání interakcí DNA s proteiny Polymerázová řetězová reakce (Polymerase chain reaction PCR) Malé
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Studijní materiály pro bioinformatickou část ViBuChu. úloha II. Jan Komárek, Gabriel Demo
 Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární genetika (Molekulární základy dědičnosti)
") Molekulární genetika (Molekulární základy dědičnosti) Struktura nukleové kyseliny Cukerná pentóza: 2-deoxy-D-ribóza D-ribóza Fosfátový zbytek: PO 4 3- Purin Pyrimidin Dusíkatá báze Adenin Guanin Tymin
Molekulární genetika (Molekulární základy dědičnosti) Struktura nukleové kyseliny Cukerná pentóza: 2-deoxy-D-ribóza D-ribóza Fosfátový zbytek: PO 4 3- Purin Pyrimidin Dusíkatá báze Adenin Guanin Tymin
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
ANALYSIS OF SERPINE1 GENE VARIABILITY IN PIGS
 ANALYSIS OF SERPINE1 GENE VARIABILITY IN PIGS Weisz F., Knoll A. Department of Animal Morphology, Physiology and Genetics, Faculty of Agronomy, Mendel University of Agriculture and Forestry in Brno, Zemedelska
ANALYSIS OF SERPINE1 GENE VARIABILITY IN PIGS Weisz F., Knoll A. Department of Animal Morphology, Physiology and Genetics, Faculty of Agronomy, Mendel University of Agriculture and Forestry in Brno, Zemedelska
Univerzita Palackého v Olomouci. Diplomová práce
 Univerzita Palackého v Olomouci Diplomová práce Olomouc 2010 Bc. Barbora Klocová Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra buněčné biologie a genetiky Mapování genů kvantitativních
Univerzita Palackého v Olomouci Diplomová práce Olomouc 2010 Bc. Barbora Klocová Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra buněčné biologie a genetiky Mapování genů kvantitativních
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární genetika (Molekulární základy dědičnosti) 0 Gen - historie 1909 Johanssen
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Molekulární genetika (Molekulární základy dědičnosti) 0 Gen - historie 1909 Johanssen
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
základní znaky živých systémů (definice života výčtem jeho vlastností) složitá organizace a řád regulace a udržování vnitřní homeostázy získávání a
 složitá organizace a řád regulace a udržování vnitřní homeostázy získávání a") definice života živý organismus je přirozeně se vyskytující sám sebe reprodukující systém, který vykonává řízené manipulace s hmotou, energií a informací základní znaky živých systémů (definice života
definice života živý organismus je přirozeně se vyskytující sám sebe reprodukující systém, který vykonává řízené manipulace s hmotou, energií a informací základní znaky živých systémů (definice života
Havarijní plán PřF UP
 Havarijní plán PřF UP v němž se nakládá s geneticky modifikovanými organismy (GMO), zpracovaný podle 20, odst. 4 zákona č. 78/2004 Sb. pro pracoviště kateder Buněčné biologie a genetiky a Oddělení molekulární
Havarijní plán PřF UP v němž se nakládá s geneticky modifikovanými organismy (GMO), zpracovaný podle 20, odst. 4 zákona č. 78/2004 Sb. pro pracoviště kateder Buněčné biologie a genetiky a Oddělení molekulární
Mutageneze vznik chyby na DNA mutagen (chemická látka / záření)
") Genotoxicita - úvod Genotoxicita: toxická látka ovlivňuje genetický materiál buňky (nukleové kyseliny) Při působení vyšších koncentrací genotoxických látek dochází k přímému úhynu buněk Nižší koncentrace
Genotoxicita - úvod Genotoxicita: toxická látka ovlivňuje genetický materiál buňky (nukleové kyseliny) Při působení vyšších koncentrací genotoxických látek dochází k přímému úhynu buněk Nižší koncentrace
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK
 ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
Molekulární genetika
 Molekulární genetika Upozornění: ukončení semestru ZÁPOČTOVÝ TEST a) Dědičnost krevně skupinových systémů (AB0, MN, Rh) b) Přepis úseku DNA do sekvence aminokyselin c) Populační genetika výpočet frekvence
Molekulární genetika Upozornění: ukončení semestru ZÁPOČTOVÝ TEST a) Dědičnost krevně skupinových systémů (AB0, MN, Rh) b) Přepis úseku DNA do sekvence aminokyselin c) Populační genetika výpočet frekvence
Biologie buňky. systém schopný udržovat se a rozmnožovat
 Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Základy genomiky. I. Úvod do bioinformatiky. Jan Hejátko
 Základy genomiky I. Úvod do bioinformatiky Jan Hejátko Masarykova univerzita, Oddělení funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky I. Zdrojová literatura ke
Základy genomiky I. Úvod do bioinformatiky Jan Hejátko Masarykova univerzita, Oddělení funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky I. Zdrojová literatura ke
Molekulární diagnostika
 Molekulární diagnostika Odry 11. 11. 2010 Michal Pohludka, Ph.D. Buňka základní jednotka živé hmoty Všechny v současnosti známé buňky se vyvinuly ze společného předka, tedy buňky, která žila asi před 3,5-3,8
Molekulární diagnostika Odry 11. 11. 2010 Michal Pohludka, Ph.D. Buňka základní jednotka živé hmoty Všechny v současnosti známé buňky se vyvinuly ze společného předka, tedy buňky, která žila asi před 3,5-3,8
Zdrojem je mrna. mrna. zpětná transkriptáza. jednořetězcová DNA. DNA polymeráza. cdna
 Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
velké fragmenty střední fragmenty malé fragmenty
 velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
velké fragmenty střední fragmenty malé fragmenty Southern 1975 Northern Western denaturace DNA hybridizace primerů (annealing) (mají délku kolem 20 bází) syntéza nové DNA termostabilní polymerázou vstup
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
MUTAGENEZE in vitro. postupy kterými se mění primární struktura DNA, především za účelem fenotypové změny
 MUTAGENEZE in vitro postupy kterými se mění primární struktura DNA, především za účelem fenotypové změny Mutace gen transkripce translace mutace normalní protein normální fenotyp mutovaný gen abnormální
MUTAGENEZE in vitro postupy kterými se mění primární struktura DNA, především za účelem fenotypové změny Mutace gen transkripce translace mutace normalní protein normální fenotyp mutovaný gen abnormální
Na rozdíl od genomiky se funkční genomika zaměřuje na dynamické procesy, jako je transkripce, translace, interakce protein - protein.
 FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
Virtuální svět genetiky 1. Translace
 (překlad) je druhým krokem exprese genetické informace a ukončuje dráhu DNA > RNA > protein. probíhá mimo jádro, v cytoplazmě na ribozómech. Výchozími látkami pro translaci je 21 standardních aminokyselin,
(překlad) je druhým krokem exprese genetické informace a ukončuje dráhu DNA > RNA > protein. probíhá mimo jádro, v cytoplazmě na ribozómech. Výchozími látkami pro translaci je 21 standardních aminokyselin,
+ Vektor. Produkce rekombinantních proteinů. Teoretický úvod. Gen. Rekombinantní DNA. Hostitelská buňka. Aplikovaná bioinformatika, Jaro 2013
 Gen + Vektor + Rekombinantní DNA Hostitelská buňka Produkce rekombinantních proteinů Teoretický úvod Aplikovaná bioinformatika, Jaro 2013 Práce s proteiny Zisk proteinů Protein je správně sbalený, aktivní,
Gen + Vektor + Rekombinantní DNA Hostitelská buňka Produkce rekombinantních proteinů Teoretický úvod Aplikovaná bioinformatika, Jaro 2013 Práce s proteiny Zisk proteinů Protein je správně sbalený, aktivní,
MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ. Agronomická fakulta BAKALÁŘSKÁ PRÁCE
 MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ Agronomická fakulta BAKALÁŘSKÁ PRÁCE BRNO 2006 Petra PŘICHYSTALOVÁ MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ Agronomická fakulta Ústav morfologie,
MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ Agronomická fakulta BAKALÁŘSKÁ PRÁCE BRNO 2006 Petra PŘICHYSTALOVÁ MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ Agronomická fakulta Ústav morfologie,
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Výuka genetiky na Přírodovědecké fakultě UK v Praze
 Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
thaliana. balky. 1. Genetická analýza a identifikace počtu genů 2. Určení DNA markerů v genetické vazbě s genem
 Praktikum z genetiky rostlin JS 2014 Genetická analýza a genetické markery 1. Genetická analýza a identifikace počtu genů odolnosti k padlí u ječmene. 2. Určení DNA markerů v genetické vazbě s genem odolnosti
Praktikum z genetiky rostlin JS 2014 Genetická analýza a genetické markery 1. Genetická analýza a identifikace počtu genů odolnosti k padlí u ječmene. 2. Určení DNA markerů v genetické vazbě s genem odolnosti
Molekulární genetika, mutace. Mendelismus
 Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Bioinformatika a výpočetní biologie KFC/BIN. I. Přehled
 Bioinformatika a výpočetní biologie KFC/BIN I. Přehled RNDr. Karel Berka, Ph.D. Univerzita Palackého v Olomouci Definice bioinformatiky (Molecular) bio informatics: bioinformatics is conceptualising biology
Bioinformatika a výpočetní biologie KFC/BIN I. Přehled RNDr. Karel Berka, Ph.D. Univerzita Palackého v Olomouci Definice bioinformatiky (Molecular) bio informatics: bioinformatics is conceptualising biology
REPREZENTACE A ZPRACOVÁNÍ GENOMICKÝCH SIGNÁLŮ REPRESENTATION AND PROCESSING OF GENOMIC SIGNALS
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
TRANSLACE - SYNTÉZA BÍLKOVIN
 TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
Amplifikační metody v molekulární diagnostice mikroorganismů. doc. RNDr. Milan Bartoš, Ph.D.
 Amplifikační metody v molekulární diagnostice mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2012 Doporučená literatura 1) Persing et al. (1993): Diagnostic Molecular
Amplifikační metody v molekulární diagnostice mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2012 Doporučená literatura 1) Persing et al. (1993): Diagnostic Molecular
Aplikovaná bioinformatika
 Aplikovaná bioinformatika Číslo aktivity: 2.V Název klíčové aktivity: Na realizaci se podílí: Implementace nových předmětů do daného studijního programu doc. RNDr. Michaela Wimmerová, Ph.D., Mgr. Josef
Aplikovaná bioinformatika Číslo aktivity: 2.V Název klíčové aktivity: Na realizaci se podílí: Implementace nových předmětů do daného studijního programu doc. RNDr. Michaela Wimmerová, Ph.D., Mgr. Josef
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS
 ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS IZOLACE FOSFOPROTEOMU A JEHO VYUŽITÍ PŘI STUDIU ÚČINKU CYTOKININŮ NA ROSTLINU Černý M., Brzobohatý B. Department
ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS IZOLACE FOSFOPROTEOMU A JEHO VYUŽITÍ PŘI STUDIU ÚČINKU CYTOKININŮ NA ROSTLINU Černý M., Brzobohatý B. Department
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Transgeneze u ptáků: očekávání vs. realita
 Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Metody používané v MB. analýza proteinů, nukleových kyselin
 Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Metody používané v MB analýza proteinů, nukleových kyselin Nukleové kyseliny analýza a manipulace Elektroforéza (délka fragmentů, čistota, kvantifikace) Restrikční štěpení (manipulace s DNA, identifikace
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
(molekulární) biologie buňky
 biologie buňky") (molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
(molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
DMPK (ZNF9) V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M
 V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M") FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Kontrola genové exprese
 Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH
 RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání
RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání